2)(超)好熱菌由来蛋白質の研究
■ 研究の背景と目的
高度好熱菌や超好熱菌が生産する蛋白質は中温生物由来蛋白質より一般に耐熱性が高い。しかし多くの場合、これらの蛋白質の耐熱化機構は明らかにされていない。 なぜなら、好熱菌由来蛋白質と中温生物由来蛋白質の三次構造を比較しても、これらの蛋白質の耐熱性の違いをもたらす構造因子を同定することは困難だからである。そこで、機能的構造的に類似しているにもかかわらず 活性の至適温度や耐熱性が大きく異なる2種類の酵素をペアとして選び、これらの性質の違いをもたらす因子を蛋白工学的手法や進化工学的手法により同定することを研究の目的とする。 このようにして蛋白質の耐熱化機構に関する理解が深められれば、作業環境に合わせて酵素の構造や機能を最適化するための設計技術を開発することも可能になる。また、超好熱始原菌(古細菌)は、 後述するように他の生物よりは原始生命体の名残を多く留めていると考えられているので、蛋白質の進化を探ることも研究の目的とする。蛋白質としては、リボヌクレアーゼH(RNase H)、サチライシン、 キヌレニンアミノトランスフェラーゼII、TBP結合蛋白質(TIP)、グリセロールキナーゼなどを用いる。また好熱菌としては、超好熱始原菌(古細菌)Thermococcus kodakarensis KOD1株を主として用いる。
・T. kodakarensis KOD1株とは?
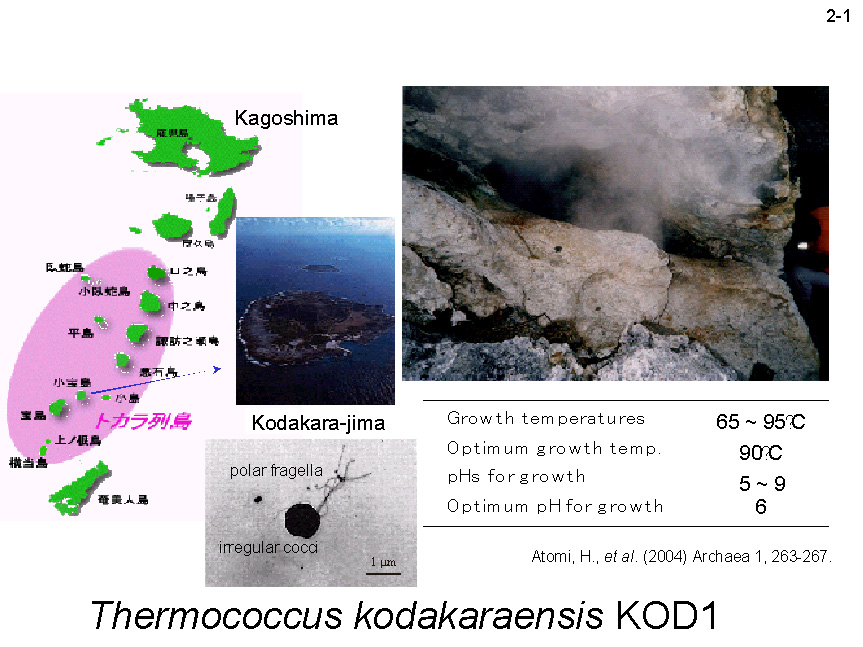
KOD1株は超好熱始原菌(古細菌)(Hyperthermophilic archaeon)の一種で、鹿児島県子宝島の硫気抗より単離された(2-1)。その生育温度範囲は60-100℃で、生育最適温度は85℃である[Atomi, H. et al. (2004) Archaea 1, 263-267]。
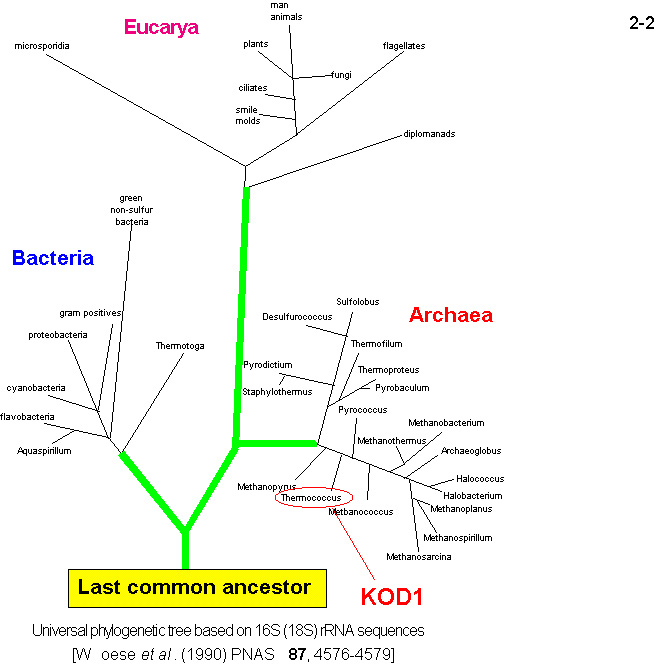
16S rRNAの配列に基づく進化系統的解析 によると[Woese, C. R. et al.
(1990) Proc. Natl. Acad. Sci. USA 87, 4576-4579]、始原菌(古細菌)(Archaea)は細菌(Bacteria)や真核生物(Eukaryotes)とは 異なるグループに分類されるが、どちらかというと真核生物と近縁である(2-2)。従って、進化的には始原菌(古細菌)と真核生物の共通の祖先がまず細菌から分かれ、ついで始原菌(古細菌)と真核生物が互いに分かれたと考えられる。
一方、超好熱始原菌(古細菌)の生育環境は原始地球環境に類似していると考えられるので、超好熱始原菌(古細菌)は、地球上に最初に誕生した原始生命体の名残を少なくとも細菌や真核生物よりは多く留めていると考えられている。
このように、超好熱始原菌(古細菌)は原始生命体と真核生物の両方の性質を合わせ持つことが期待されるので、KOD1株は生命の進化をたどる上でも貴重な生物資源である。
主な研究テーマ
(1)KOD1株由来RNase H2(Tk-RNase H2)の耐熱化機構の解析
{kind=link}
{kind=link}