(1)KOD1 株由来 RNase H2(Tk-RNase H2)の耐熱化機構の解析
T. kodakarensis KOD1由来RNase H2(Tk-RNase H2) は type
2 RNase H で、type 1 RNase H の大腸菌RNase H1( Ec-RNase H1) とは一次構造的に異なる(1-2-2)[Haruki,
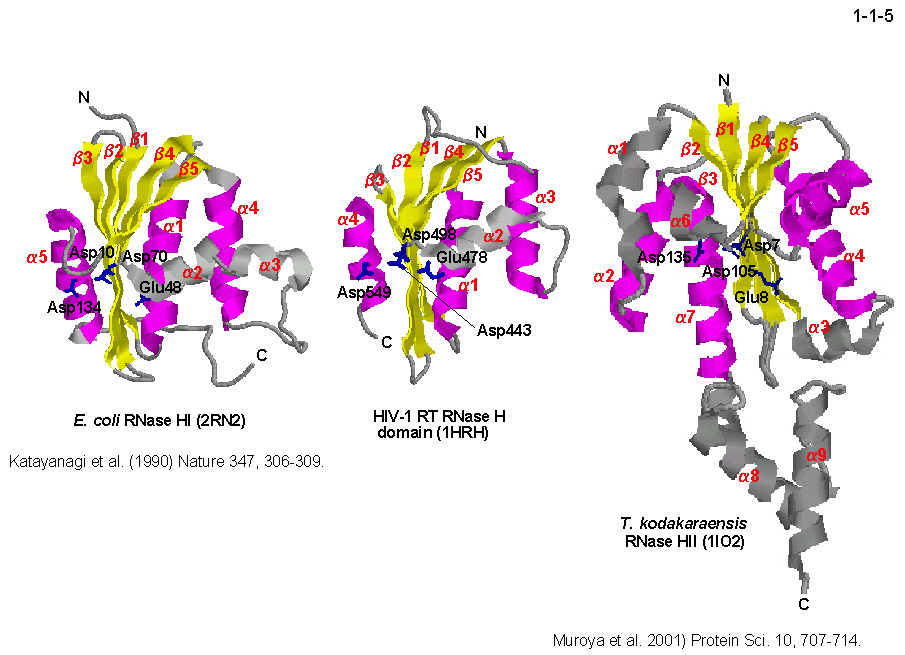
M. et al. (1998) J. Bacteriol. 180, 6207-6214]。 しかし、その三次構造は Ec-RNase H1と似ている(1-1-5)[Muroya,
A. et al. (2001) Protein Sci. 10,
707-714]。Tk-RNase H2 も Ec-RNase
H1もRNase H-fold と呼ばれる構造モチーフを持っており、 Tk-RNase H2 の活性部位を構成する4残基の酸性アミノ酸(Asp7、Glu8、Asp105、Asp135)の空間配置は Ec-RNase
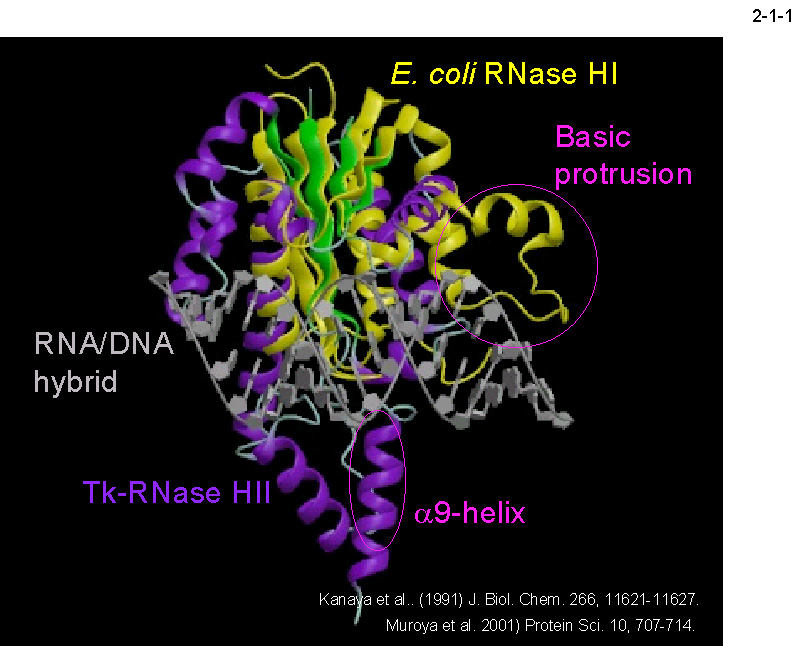
H1 の4残基の酸性アミノ酸(Asp10、Glu48、Asp70、Asp134)の配置とよく似ている。従って、 これらの酵素の触媒反応機構はよく似ていると考えられる。しかし、Ec-RNase H1 の場合は分子中央部に位置する塩基性突出部位と呼ばれるドメインが基質結合に関与するのに対して[Kanaya, S. et al. (1991) J. Biol. Chem. 266, 11621-11627; Haruki, M. et al. (1997) J. Biol. Chem. 272,
22015-22022]、 Tk-RNase
H2 の場合は C 末端ドメインのα9 ヘリックスが基質結合に関与する(2-1-1)[Muroya,
A. et al. (2001) Protein Sci. 10,
707-714]。
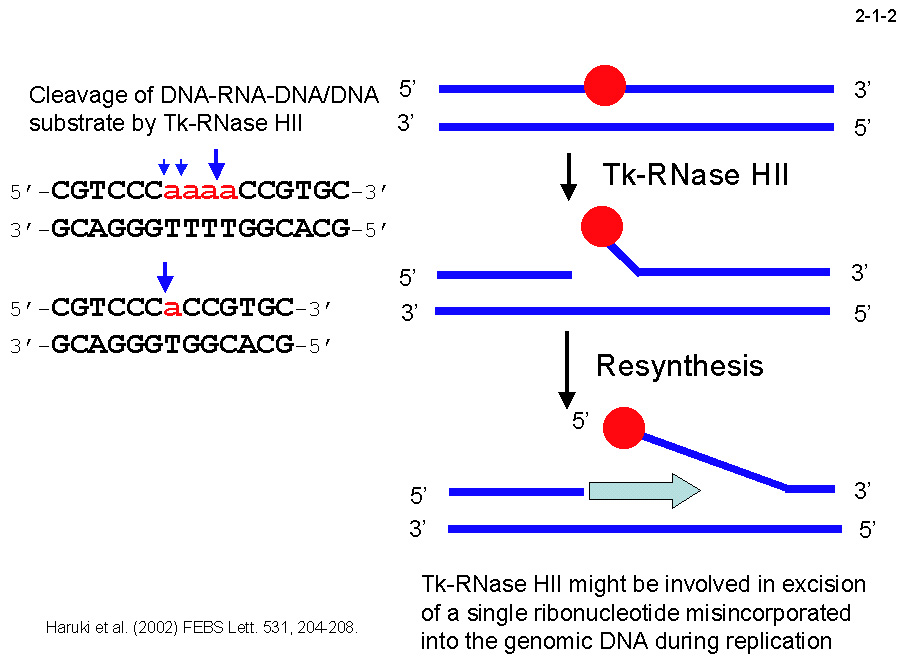
Tk-RNase H2 は 228 残基から成りモノマーとして働く。他の RNase H 同様二価金属イオンを活性に必要とするが、 Ec-RNase H2やEc-RNase H2が Mg2+と Mn2+存在下でのみ活性を示すのに対して、 Tk-RNase H2 はこの二種に加え、Co2+、Ni2+存在下でも活性を示す[Haruki, M. et al. (1998) J. Bacteriol. 180, 6207-6214]。つまり、Tk-RNase H2 は前二者に比べて幅広い金属イオン特異性を示す。 Tk-RNase H2 は DNA-RNA-DNA/DNA 基質の RNA-DNA 連結部をリボヌクレオチドが1残基だけ残る位置で切断する(2-1-2)。 また、DNA-RNA 連結部も切断する。これは、この基質が RNA を1残基だけ含む場合も同じである。つまり Tk-RNase H2 は、 RNA を1残基だけ含む DNA-RNA-DNA/DNA 基質をこの RNA の 5’側で切断する。従って、 Tk-RNase
H2 は複製中に誤って取込まれた RNA の除去に関与すると考えられる(2-1-2)[Haruki, M. et al.
(2002) FEBS Lett. 531,
204-208]。なお、 Ec-RNase
H1は DNA-RNA-DNA/DNA 基質の DNA-RNA 連結部を切断しないし、RNA-DNA 連結部もリボヌクレオチドが2つ以上残る位置で切断する。
従って Ec-RNase H1 は、RNA を1残基だけ含む DNA-RNA-DNA/DNA 基質を切断しない。このような基質切断部位の違いは、 GRGモチーフと保存Tyr残基の有無に起因する(GRGモチーフとTyrはRNase H2にだけ保存されている)[Rychlik, M. P. et al. (2010) Mol. Cell 40, 658-670]。
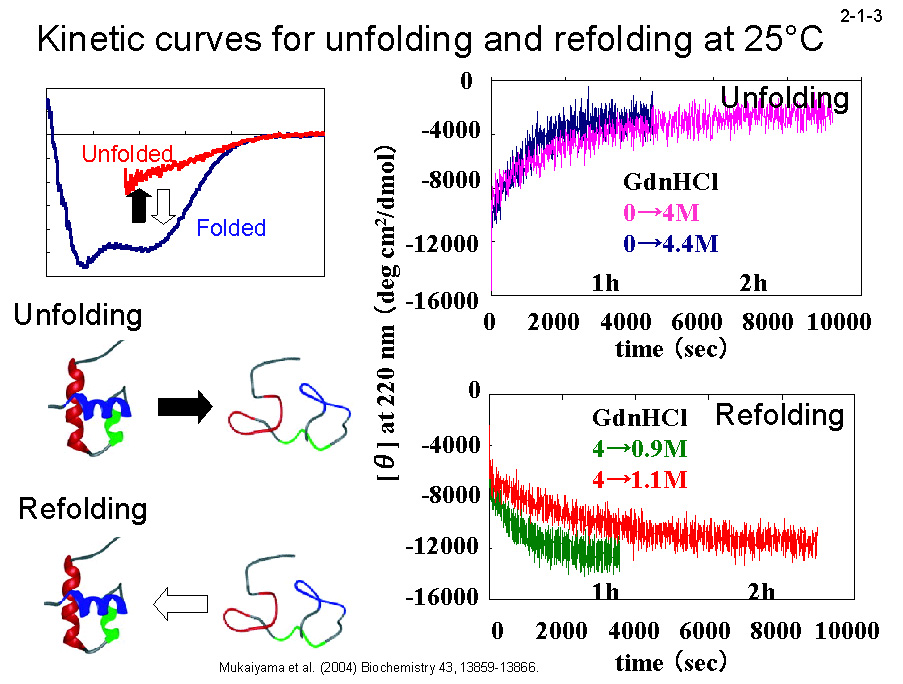
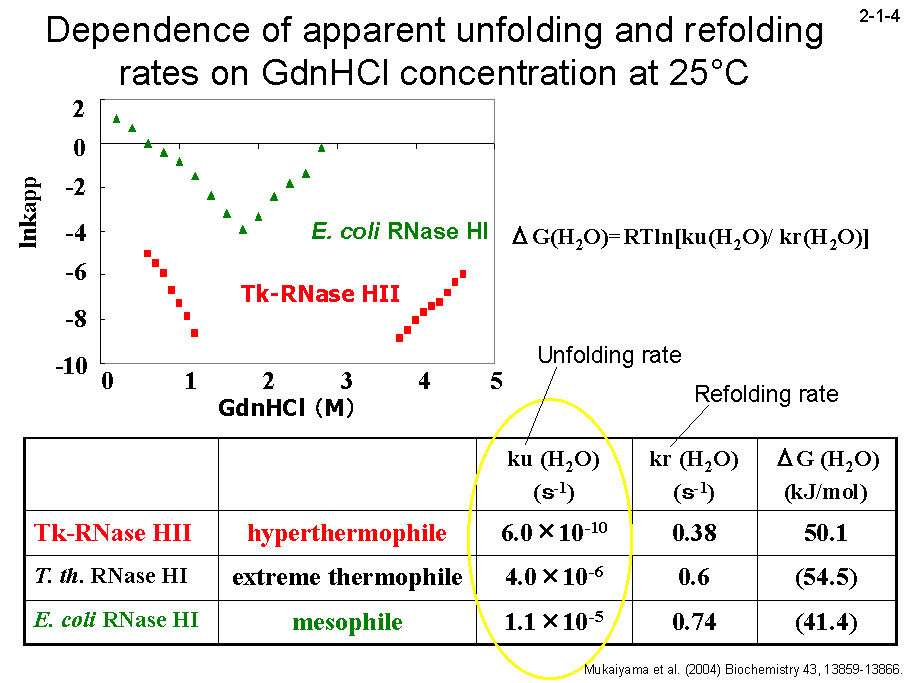
Tk-RNase H2 は 90℃ で1時間加熱してもほとんど失活しない[Haruki, M. et al. (1998) J. Bacteriol. 180, 6207-6214]。高度好熱菌由来 RNase H1(Tt-RNase H1)も同条件で失活しない。 一方、Ec-RNase H1 は 50℃ でも半減期約 15 分で失活する[Haruki, M. et al. (1998) J. Bacteriol. 180, 6207-6214]。CD を用いて解析した Tk-RNase H2 の GdnHCl 変性速度(2-1-3)は、 Tt-RNase H1 や Ec-RNaseH1 より著しく遅い(2-1-4)[Mukaiyama, A. et al.
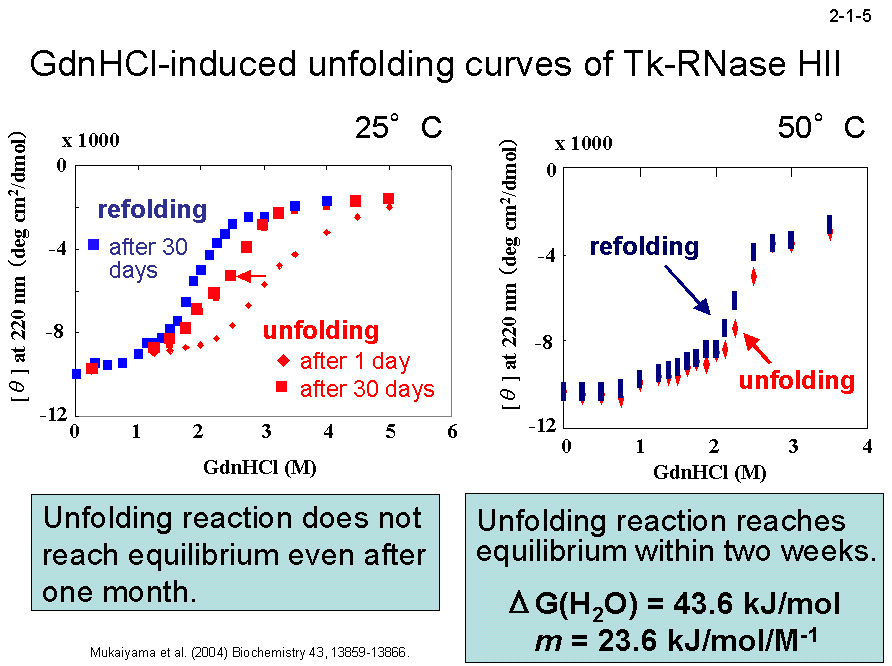
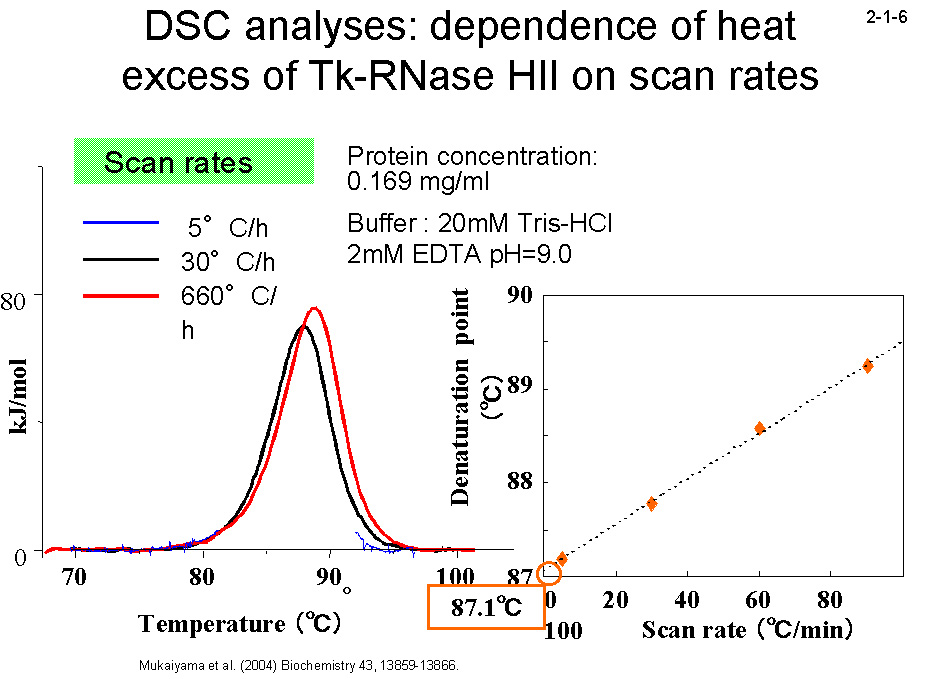
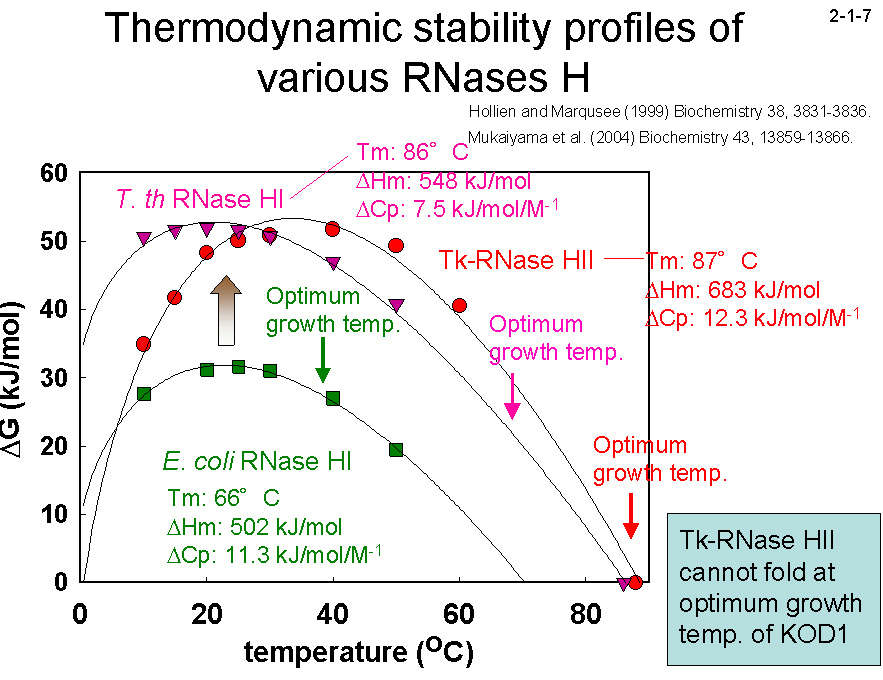
(2004) Biochemistry 43, 13859-13866]。一方、復元速度に関しては、3つの蛋白質間に大きな違いは見られない(2-1-4)。 そのために、25℃ では変性反応と復元反応は1ヶ月経過しても平衡に達しない(2-1-5)。DSC のピーク(変性温度)も昇速度に比例して上昇する(2-1-6)。 また、様々な温度で変性速度実験を行い、温度とΔG の関係を求めると、Tk-RNase H2 の曲線は Tt-RNase
H1 に比べて、高温領域に平行移動して、その安定性は 40℃ 付近で最大になる(2-1-7)。
ただし、変性温度は 87℃ とあまり変わらない。Ec-RNaseH1 と比べた場合は、Tk-RNase H2 の曲線は高ΔG 値領域にも大きく平行移動している。従って、 Tk-RNase Hは Tt-RNase H1 や Ec-RNase H1 より平衡論的にも速度論的にも安定化している。一見同じように安定であると思われている好熱菌由来蛋白質同士でも、その安定化メカニズム、
特に速度論的安定性に違いがあるのは興味深い。また、Tk-RNase H2 の変性温度が KOD1 株の最適生育温度より低いことも興味深い。おそらくKOD1の細胞内にはTk-RNase H2 の安定化に寄与する因子があり、そのためにTk-RNase H2 は90℃ 以上の高温でも細胞内では 安定に存在して機能していると考えられる。
なお、原核生物由来RNase HはRNase HI、RNase HIIと命名されてきたが[Itaya, M. (1990) Proc. Natl. Acad. Sci. USA 87, 8587-8591]、最近は真核生物由来RNase H同様、RNase H1、RNase H2が用いられる。従って、本文中ではRNase H1、RNase H2を用いた。各参考図ではRNase HI、RNase HIIのままとなっている。
<2006年以後の研究>
1)Tk-RNase H2の活性部位変異体の酵素活性、および安定性測定を行い、活性部位の構造安定性への寄与を調べた。その結果、測定したすべての活性部位変異体はその酵素活性が野生型に比べ減少しているのに対して、構造安定性は50℃でのGdnHCl変性、熱変性いずれの実験においても野生型より安定であることがわかった[Mukaiyama, A. et al. (2006) Biochemistry 45, 12673-12679]。つまり、Tk-RNase H2の活性部位を構成する4つの酸性残基はいずれも構造安定性の観点から見れば至適でないことが示された。また、野生型酵素と比べて酵素活性が著しく減少する変異体ほど、大きく安定性が増加するという相関関係が確かめられた。これらの結果は、Tk-RNase H2は機能を獲得するために構造安定性を犠牲にしていることを示唆する。
2)アミンの一種であるトリメチルアミン-N-オキシド (TMAO)が溶媒に存在することでTk-RNase H2の構造安定性が安定化されるのかを調べた。その結果、熱変性および変性剤変性のいずれにおいてもTk-RNase H2がTMAOによって安定化することが示された[Mukaiyama, A. et al. (2008) Proteins 71, 110-118]。さらに、GdnHCl変性の速度実験からこのようなTMAOによる安定性の増加は変性速度がより遅く、復元速度がより速くなることに起因していることが明らかとなった。また、TMAOによる安定化はTk-RNase H2の天然状態および変性状態のコンフォメーションに影響をおよぼさずに起こっていることがそれぞれのCDスペクトルおよび構造安定性の熱力学的解析から確認された。これらの結果は、実際の生体内では様々な低分子化合物がTk-RNase H2はじめ細胞質蛋白質の構造安定性に影響を与えていることを示唆している。
3)Tk-RNase H2の嵩高い疎水性残基をより小さな疎水性残基に系統的に置換した変異体を9種類構築し、これらの変異体の構造安定性やフォールディングを解析することにより、内部に埋もれた疎水性残基はTk-RNase H2の遅い変性反応に寄与することを明らかにした[Dong, H. et al. (2008) J. Mol. Biol. 378, 264-272]。
4)αヘリクッスN末端領域のプロリン残基が、超好熱古細菌由来タンパク質の熱安定性や変性、復元機構にどのような影響をおよぼすか調べた。その結果、αヘリクッスN末端領域のプロリン残基は、非常に高い安定性を有するTk-RNase H2においても中温生物由来タンパク質における場合と同様に加算的な安定性への寄与を示した[Takano, K. et al. (2009) J. Biochem. 145, 79-85]。特に、柔軟性の高い領域に存在することでそのタンパク質の安定性を効果的に上げていた。しかし、Tk-RNase H2の遅い変性反応には疎水性相互作用のような大きな影響を与えなかった。これは、プロリン残基による安定化は主に変性状態のタンパク質に寄与する影響であって、天然構造にあまり影響しないためと考えられる。これにより、天然状態の疎水性相互作用や水素結合などのエンタルピックな影響は遅い変性反応に強い影響を与えるが、変性状態におけるプロリン残基やジスルフィド結合等のエントロピックな影響は遅い変性反応にあまり影響を及ぼさないことが示唆された。さらに、αへリックスN末端領域のプロリン残基は古細菌由来Tk-RNase H2と細菌由来Tm-RNase
H2の変性速度が大きく異なる原因ではなかった。このことから、αへリックスN末端領域のプロリン残基はそのタンパク質の進化的背景に関係なく、普遍的にタンパク質の安定化に関与していることがわかった。
5)Tk-RNase H2と Tt-RNase H1はどちらも安定なRNase Hであり、立体構造も類似しているにも関わらず、その速度論的安定性は大きく異なっている。その違いが、RNase Hの進化的起源の違いによるのか宿主の進化的起源の違いによるのかを明らかにする目的で、超好熱細菌Thermotoga maritima由来Tm-RNase H2、超好熱細菌Aquifex aeolicus由来Aa-RNase H2、超好熱古細菌Sulfolobus tokodaii由来Sto-RNase H1の安定性を解析し、Tk-RNase H2、Tt-RNase H1、Ec-RNase H1の安定性と比較した。その結果、細菌由来のTm-RNase H2とAa-RNase H2の変性速度は細菌由来のTt-RNase H1やEc-RNase H1と同様速く、古細菌由来のSto-RNase H1の変性速度は古細菌由来のTk-RNase H2同様遅いことがわかった[Okada, J. et al. (2010) BMC Evol. Biol. 10, 207]。これらの結果は、超好熱菌由来RNase Hの速度論的安定性の違いは、RNase Hの進化的起源(type 1、type 2)ではなく、宿主の進化的起源(細菌、古細菌)の違いによることを示唆する。
6)遅い変性反応を示す超好熱古細菌由来Tk-RNase H2と遅い変性反応を示さない超好熱細菌由来Tm-RNase H2の変性過程を、超安定なプロテアーゼである超好熱菌由来Tk-subtilisinを用いて追跡するという新しい手法(pulse proteolysis)で解析した。その結果、Tk-RNase H2の変性過程は、初期段階でTk-subtilisinによる切断を受けやすい中間状態に移行し、次に部分的にTk-subtilisinに切断される中間状態に移行してからTk-subtilisin耐性を示す中間状態に移行する、多数の中間状態を介した多状態反応であることが確認できた[Okada, J. et al. (2012) Biochemistry 51, 9178-9191]。そして、Tk-RNase H2の遅い変性反応にはN末側領域が重要な役割を担い、C末端領域は復元反応と構造安定性に寄与していることが示唆された。一方、超好熱細菌由来Tm-RNase H2の変性過程は中間体を介さない二状態反応を示した。このことは、古細菌由来Tk-RNase H2と細菌由来Tm-RNase H2は、その宿主の進化的背景に依存した異なる変性機構を有していることを示唆している。さらに、本実験手法がタンパク質の変性過程を解析する手段として有効であることを示した。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}