(2)KOD1 株由来サチライシン(Tk-subtilisin、Tk-SP)の研究
(2−1)Tk-subtilisin
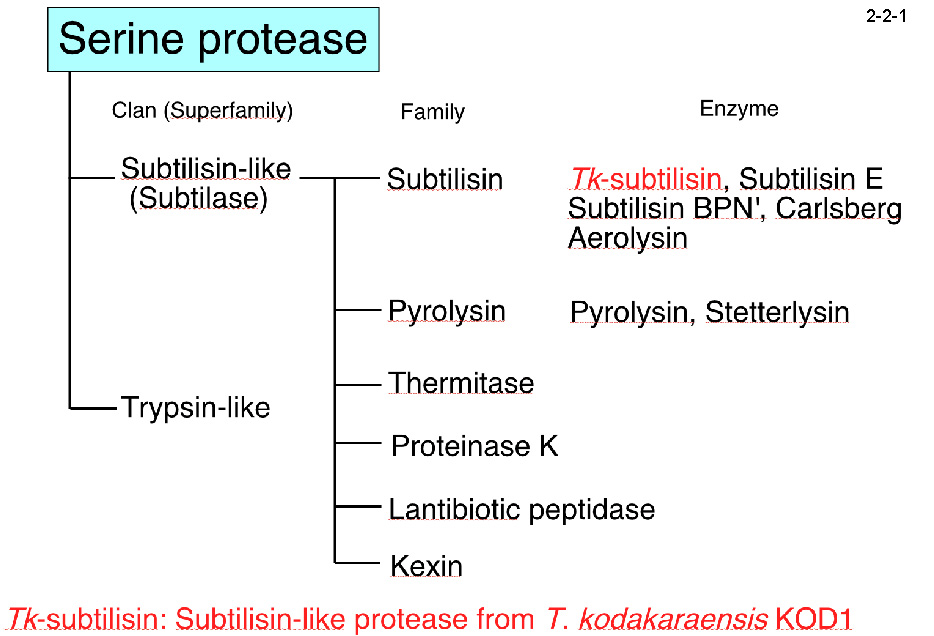
Tk-subtilisin はセリンプロテアーゼの一種で subtilisin E、subtilisin BPN、subtilisin
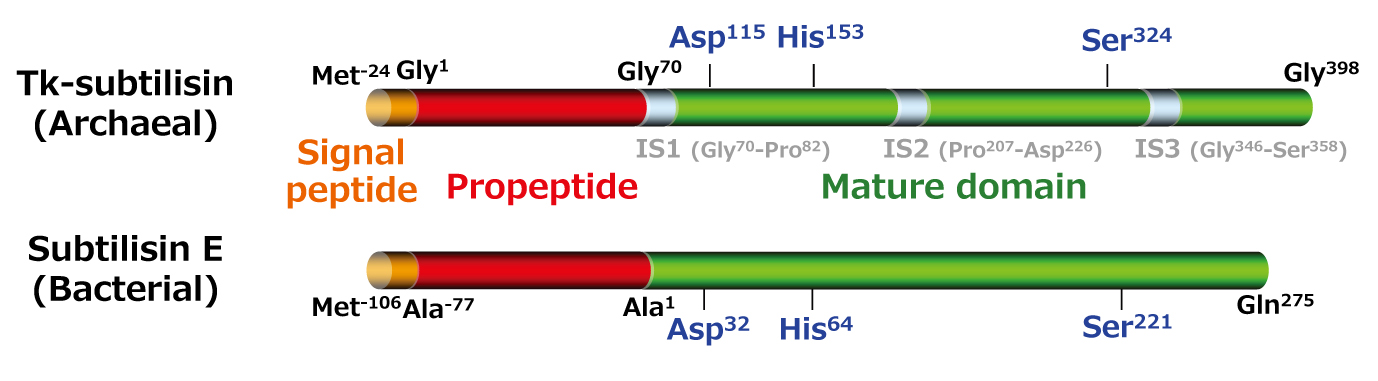
Carlsberg などと高い相同性を有する(アミノ酸配列の同一性は43-44%)(2-2-1)。 subtilisin は広く工業的に用いられる酵素であり、高い熱安定性を持つ Tk-subtilisin の研究は基礎的な面だけでなく工業的な面からも意義深い。本酵素は他の subtilisin 同様、分泌シグナルであるプレ配列[Met(-24)-Ala(-1)]、 分子内シャペロンとしても阻害剤としても働くプロペプチド(Gly1-Leu69)、そしてプロテアーゼ活性をもつ成熟体(Gly70-Gly398)から成る[Pulido, M. et al. (2006) Appl. Environ. Microbiol.
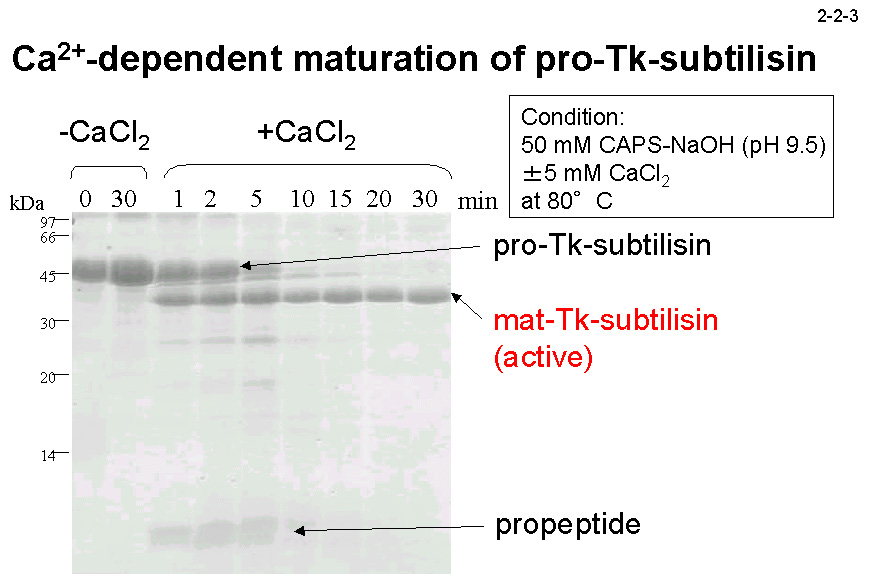
72, 4154-4162](2-2-2)。本酵素は、他の subtilisin 同様、 プロペプチドを含むプロ体(Pro-Tk-subtilisin)として培地中に分泌された後、プロペプチドが切断されて活性型の成熟体になる(2-2-3)。ただし、成熟化には Ca2+を必要とする。
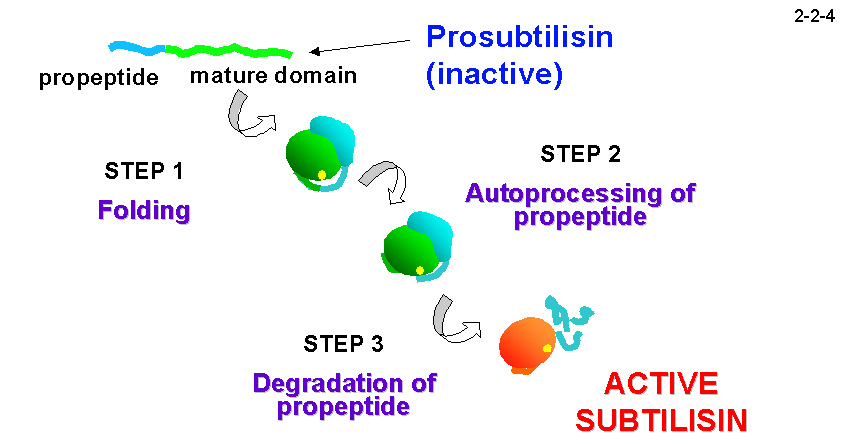
Subtilisin E などを用いたこれまでの研究から、分泌後の pro-subtilisin の成熟化は、1)プロペプチドの補助による成熟体の折り畳み、2)プロペプチドと成熟体の間のペプチド結合の自己切断、
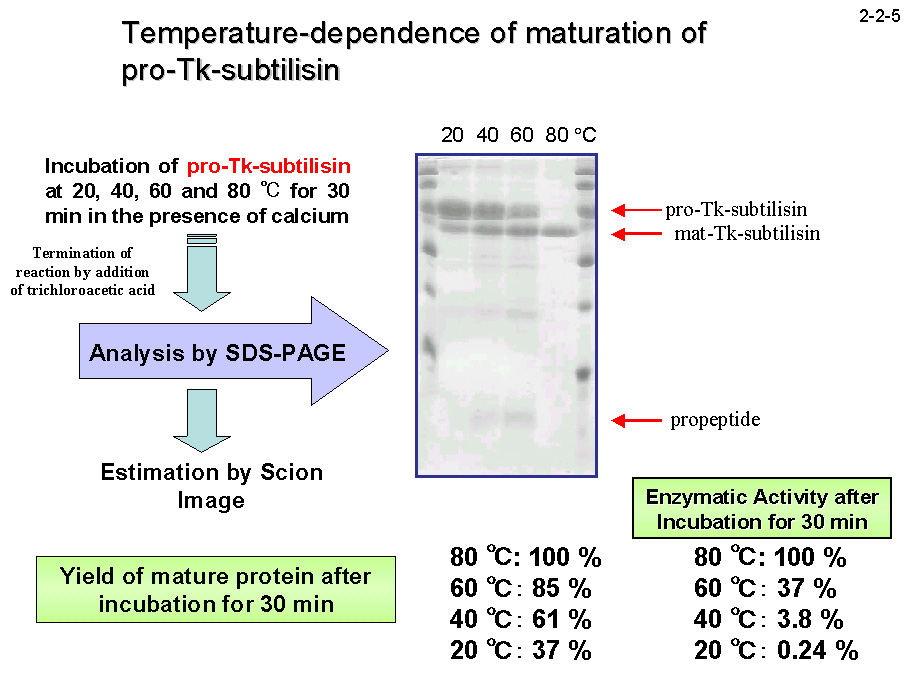
3)活性部位を塞ぐことにより活性を阻害しているプロペプチドの分解除去、の3段階のステップで進むと考えられている(2-2-4)。他の subtilisin では、ステップ1)において、 シャペロンであるプロペプチドがない場合成熟体は正しく折り畳まれず活性を示さないことから、プロペプチドはsubtilisinの成熟体が正しい構造を形成するのに不可欠であるとされている。しかし、Tk-subtilisin はプロペプチドを持たない成熟体として発現させても活性を示すので、 Tk-subtilisin のプロペプチドは成熟体の構造形成に必ずしも必須ではないと考えられる。また、Pro-Tk-subtilisin の成熟化を 40 ℃ 以下の低温で行うと、ステップ2)で停止した状態の中間体、
つまり成熟体から切り離されたプロペプチドが分解されない状態の中間体、が得られるが、この中間体は活性を示さない(2-2-5)。従って、自己切断されたプロペプチドは成熟体から解離せず、その活性を阻害する。実際、
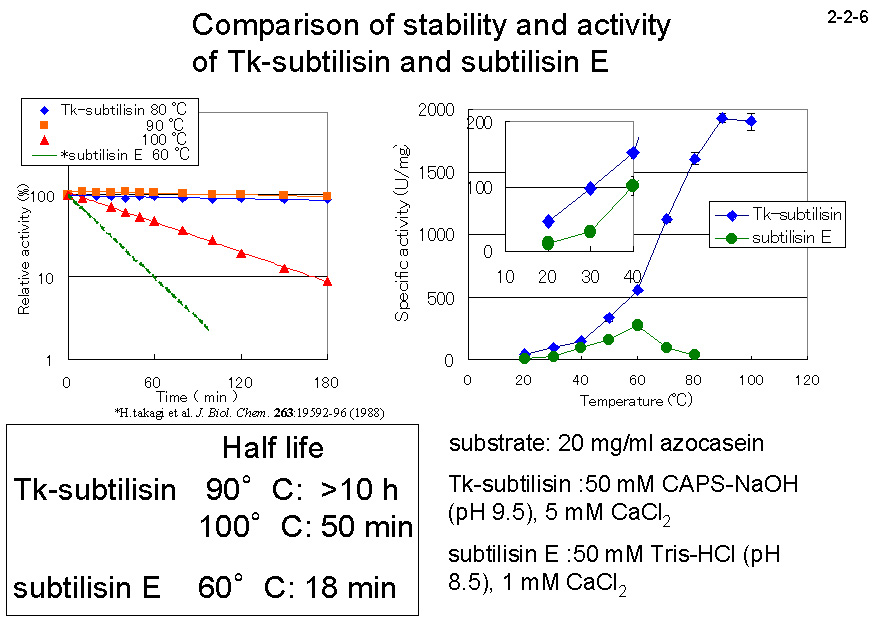
大腸菌を用いて大量発現させた Tk-subtilisin のプロペプチドは、成熟体の活性を強く阻害する。Tk-subtilisin と中温菌由来の subtilisin E の安定性や活性を比較すると、Tk-subtilisin は高い耐熱性を示す。
また、超好熱菌由来の酵素でありながら高温域のみならず中温域でも高い活性を有し、20 ℃ での活性は中温菌由来の subtilisin
E より 3.6 倍も高い(2-2-6)。 Pro-Tk-subtilisin は低温ではプロペプチドの阻害が強すぎるため成熟化しにくいが、高温ではすみやかに成熟化する。従って、Tk-subtilisinの成熟化機構は、フォールディングにプロペプチドではなくCa2+を必要とする、成熟化に高温を必要とするという点でSubtilisin Eなどの成熟化機構とは異なる [Pulido, M. et al. (2006)
Appl. Environ. Microbiol.
72, 4154-4162]。
<2007年以後の研究>
1)活性中心変異体のプロ体(Pro-S324A)および自己切断後のプロペプチドと成熟体の複合体(Tk-propeptide:S324C-subtilisin
complex)、プロペプチド分解後の成熟体(MIP-Tk-subtilisin)の結晶構造を決定した。その結果、Tk-subtilisinの成熟体にはCa2+が7個結合すること、その構造はCa2+が結合することにより大きく変化すること、成熟体の構造形成はプロペプチドの自己切断前に完了することを明らかにした[Tanaka, S. et
al. (2007) J. Biol. Chem. 282, 8246-8255;Tanaka, S. et al. (2007) J. Mol. Biol. 372, 1055-1069]。また、Tk-subtilisinの活性中心変異体の結晶構造とフォールディング速度を解析することにより、Tk-subtilisinのプロペプチドは成熟体のフォールディングを促進する働きはあるものの、フォールディングに不可欠ではないことを明らかにした[Tanaka, S. et al. (2008) FEBS Lett. 582, 3875-3878]。
3)Tk-subtilisinが高温でしか成熟化しないのは、プロペプチドが構造的に極めて安定なためであり、プロペプチドが不安定化すると成熟化の温度は低下することを、進化工学的手法を用いて明らかにした[Pulido, M. et al. (2007) Protein Eng. Des. Sel. 20, 143-153]。また、成熟化の温度が低下した変異体の結晶構造を解析することにより、プロペプチドが不安定化して分解されやすくなったのは、コア領域に歪みが生じたためであることを明らかにした[Pulido, M. et al. (2007) J. Mol. Biol.
374, 1359-1373]。
4)7個のCa2+のうち4個(Ca2-Ca5)が結合するCa2+結合ループを削除するとTk-subtilisin はCa2+存在下でもフォールディングしなくなることを明らかにした。また、Ca2あるいはCa3が結合できなくなるような変異を導入するとTk-subtilisinのフォールディング速度が大きく低下することを明らかにした。いずれの場合もTk-subtilisinの安定性に変化はなかった。以上の結果、Ca2+結合ループにCa2+が結合すると、Tk-subtilisinのコア領域(αβαサブ構造)の構造形成が誘導されることを明らかにした[Takeuchi, Y. et al. (2009) Biochemistry 48, 10637-10643]。
5)プロペプチドによるTk-subtilisinのフォールディング促進機構を明らかにする目的で、プロペプチド変異体を各種構築し、そのフォールディング促進能を解析した。その結果、Tk-subtilisinのコア領域(αβαサブ構造)と相互作用するループ領域が、プロペプチドによるフォールディングの開始に必要であることを明らかにした[Tanaka, S. et al. (2009) J. Mol. Biol. 394, 306–319]。この結果は、Tk-subtilisinがフォールディングのための高いエネルギー障壁を2つの異なる方法(Ca2+の結合とプロペプチドとの相互作用)で下げていることを示唆しており興味深い。
6)7個のCa2+のうちCa7、Ca6、Ca1がそれぞれ結合できなくなる変異を導入すると、Tk-subtilisinの安定性がこの順に大きく低下することを明らかにした[Uehara, R. et al. (2012) Biochemistry 51, 5369–5378]。従って、Tk-subtilisinはこれらのCa2+の結合によって高い安定性を獲得したと考えられる。いずれの場合もTk-subtilisinのフォールディング速度に変化はなかった。この結果は、Tk-subtilisinがフォールディングと安定性にCa2+を使い分けていることを示しており興味深い。
7)プロペプチドと成熟体の間の挿入配列IS1を除去すると、プロ体の構造が著しく不安定化することを明らかにした[Uehara, R. et al. (2012) Extremophiles 16, 841-851]。しかし、プロ体が変性するような高温でない限り、成熟化速度に大きな変化は見られず、成熟体の活性や安定性にも変化は見られなかった。以上の結果、IS1はTk-subtilisinの成熟化において、プロペプチド切断前の構造の安定化に寄与しており、その成熟化を高温環境に適応させる機能を持つと考えられた。
8)触媒ドメインのS1サイトに結合するプロペプチドのC末端のLeuをProに置換すると、プロペプチドと成熟体の間のペプチド結合は自己切断されにくくなるが、自己切断によって生じたプロペプチドがTk-subtilisinによって分解されやすく速度が向上するため、結果的に成熟化が加速されることを明らかにした。触媒ドメインのフォールディング速度に変化は見られなかった。一方、この変異によりプロペプチドの阻害能やTk-subtilisinへの親和性が低下した。以上の結果、触媒ドメインとの親和性を低下させるプロペプチドC末端への変異導入が、Tk-subtilisinの成熟化速度を促進することを明らかにした[Uehara, R. et al. (2012) in
press]。
(2−2)Tk-SP
Tk-SP はsubtilisinファミリーのセリンプロテアーゼで subtilisin E、subtilisin BPN、subtilisin Carlsberg などと約40%、Tk-subtilisinと31%のアミノ酸配列の相同性を示す。 Tk-subtilisin 同様、高い耐熱性を示し、界面活性剤や変性剤などに対しても高い安定性を示すので、産業酵素としての利用が期待されている。特に、Tk-subtilisinと異なりCa2+イオンをフォールディングに必要としないので、キレート剤に対しても比較的高い安定性を示す。本酵素は他の subtilisin 同様、分泌シグナルであるプレ配列[Met(-23)-Ala(-1)]、阻害剤として働くプロペプチド(Ala1-Ala113)、そしてプロテアーゼ活性をもつ成熟体(Val114-Gly640)から成る。成熟体はサチライシンドメイン(Val114-Tyr421)、βジェリーロールドメイン(Ala422-Val539)、C末端ドメイン(Asp540-Gly640)から成る。本酵素は、他の subtilisin 同様、 プロペプチドを含むプロ体(Pro-Tk-SP)として培地中に分泌された後、プロペプチドが切断されて活性型の成熟体になる。この成熟化には Ca2+もプロペプチドも必要としない。ただし、C末端ドメインはCa2+がないと不安定で自己分解されやすいので、Tk-SPあるいはその前駆体をCa2+非存在下で熱処理(80℃、30分)するとC末端ドメインが完全に分解除去されたTk-SP*(Val114-Val539)が生成する。Tk-SP*はTk-SPよりやや不安定で活性もやや低いが、Tk-subtilisin同様、高い耐熱性を示す。
<これまでの研究>
1)Pro-Tk-SPは大腸菌菌体内に可溶性タンパク質として蓄積するが、培養中、精製中に成熟化が進行し、最終的にTk-SP*として得られることを明らかにした。また、Tk-SP*の至適温度が100℃、100℃での半減期が100分であること、5%SDSや8M尿素で処理しても変性しないこと、EDTA処理すると安定性は低下するものの80℃でも安定であることを明らかにした[Foophow, T. et al. (2010)
Protein Eng. Des. Sel. 23, 347-3553]。
2)Pro-Tk-SPの活性中心変異体でC末端ドメインの欠如したPro-S359A*の結晶構造を決定した[Foophow, T. et al. (2010) J. Mol. Biol. 400, 865-877]。その結果、Pro-S359A*はプロペプチド、subtilisinドメイン、βジェリードメインから成ることを明らかにした。また、βジェリードメイン以外の構造はPro-Tk-subtilisinの活性中心変異体の構造と良く似ていること、2個のCa2+がβジェリードメインに結合していることを明らかにした。さらに、βジェリードメインがTk-SPの高い耐熱性に寄与すること、この寄与はCa2+が結合した時だけ見られることを明らかにした。
3)微弱な活性を示すPro-Tk-SPの活性中心変異体Pro-S359Cおよびその誘導体の活性や安定性を解析することにより、βジェリードメインは活性には必要ないこと、C末端ドメインはCa2+存在下では自己分解されにくいこと、C末端ドメインは活性や安定性に関与することを明らかにした[Sinsereekul, N. et al. (2011) FEBS J. 278, 1901-1911]。以上の結果、Tk-SP*ではなくTk-SPが成熟体であることを明らかにした。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}