(3)その他の超好熱菌由来蛋白質の研究
T. kodakaraensis KOD1 株由来グリセロールキナーゼ(Tk-GK)の耐熱化機構の解析、T. kodakaraensis KOD1 株由来 TBP 結合蛋白質(Tk-TIP)の機能解析と結晶構造解析、
超好熱始原菌(古細菌)Pyrococcus horikoshii OT3株由来キヌレニンアミノトランスフェラーゼ II(Ph-KAT II)の結晶構造解析、超好熱性細菌Thermotoga malitimaおよびAquifex aeolicus由来 RNases H
(Tm-RNases HI, HII、Aa-RNases HII, HIII)の機能解析と結晶化、などを行っている。
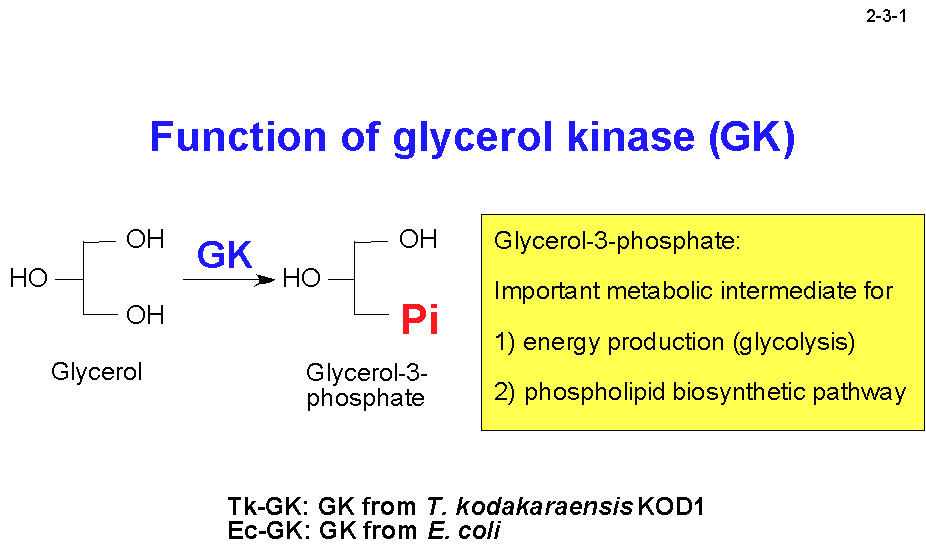
KOD1 株由来グリセロールキナーゼ(Tk-GK)は ATP 依存性の酵素で、グリセロールをリン酸化してエネルギー代謝系およびリン脂質合成系における重要な中間代謝物であるグリセロール3リン酸を生成する(2-3-1)。
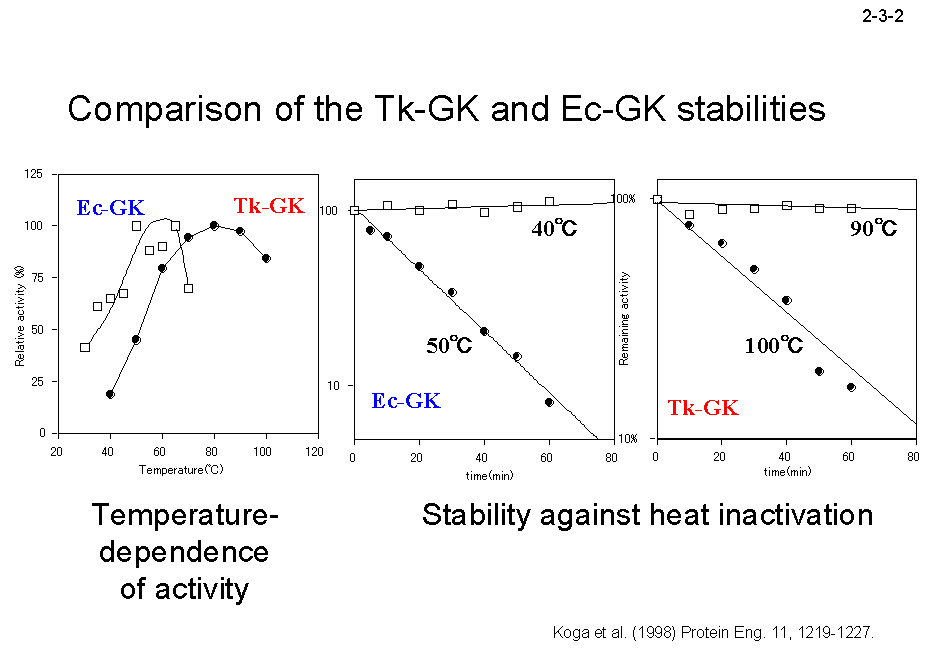
Tk-GK は 497 アミノ酸残基から成り、大腸菌由来 GK (Ec-GK)と一次構造上 57% の同一性を示す。しかし、Tk-GK は Ec-GK(4量体)とは異なり2量体として働く。また、Tk-GK は Ec-GK より広い2価金属イオン特異性を示し、
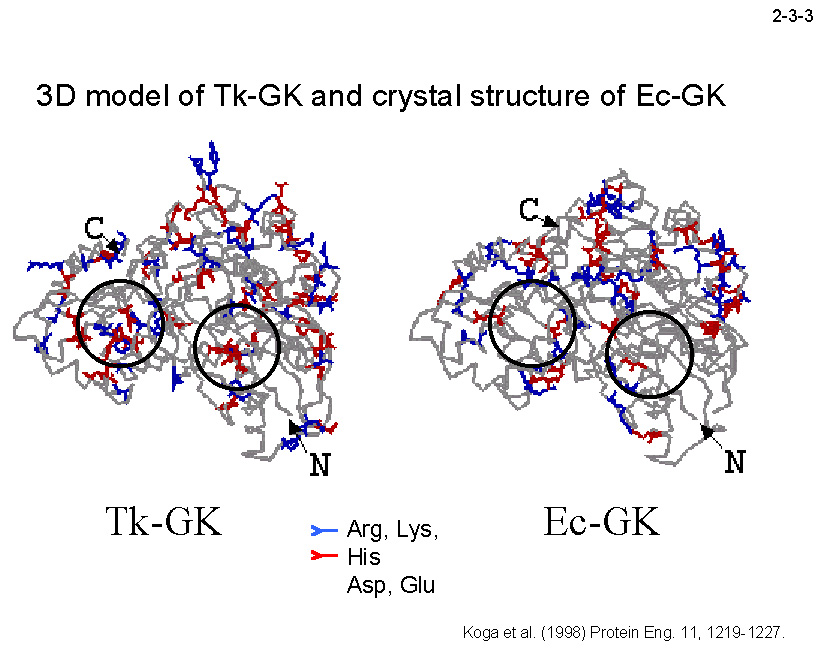
Mg2+だけでなく Co2+、Zn2+、Mn2+、Ni2+存在下でも活性を示す。さらに、Tk-GKはEc-GKより著しく耐熱性が高い(2-3-2)。Tk-GK の三次構造モデルを Ec-GK の結晶構造と比較することにより、
分子表面のイオンペアネットワークの増加やループ部分の Pro の増加が Tk-GK の耐熱化に寄与することが示唆されている(2-3-3)。また、DNA シャフリング法により、
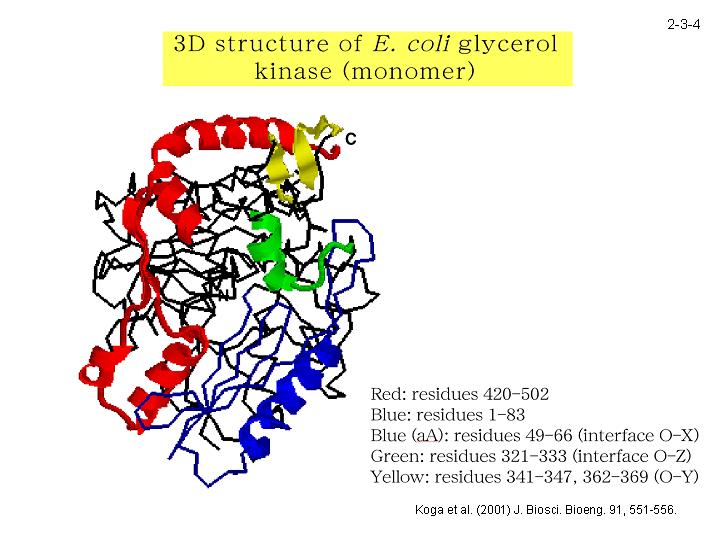
Tk-GK と Ec-GK の安定性の違いをもたらすアミノ酸置換は C 末端領域に局在していることを明らかにしている(2-3-4)。今後、このようなアミノ酸置換の同定と Tk-GK の結晶化を行う予定である。
KOD1株由来 TBP 結合蛋白質(Tk-TIP, TIP: TBP-interacting protein)は TBP を固定化したアフィニティカラムを用いて KOD1 株の細胞抽出液から単離されている。
その部分配列に基づいて設計したプライマーを用いて遺伝子もクローニングされ、組換え蛋白質の大量生産系も構築されている。興味深いことに、本蛋白質のホモログは KOD1 株と近縁の一部の超好熱始原菌(古細菌)にしか存在しない。
本蛋白質は 224 アミノ酸残基から成り2量体として存在する。ゲルシフトアッセイにより、本蛋白質は TBP と結合することにより TBP と DNA の相互作用を阻害することや、TBP/TFB/DNA と複合体を形成することを明らかしている。
従って、本蛋白質は転写に関与すると考えられる。本学工学研究科甲斐研との共同研究として本蛋白質の結晶化にも成功しており現在構造の精密化をすすめている。
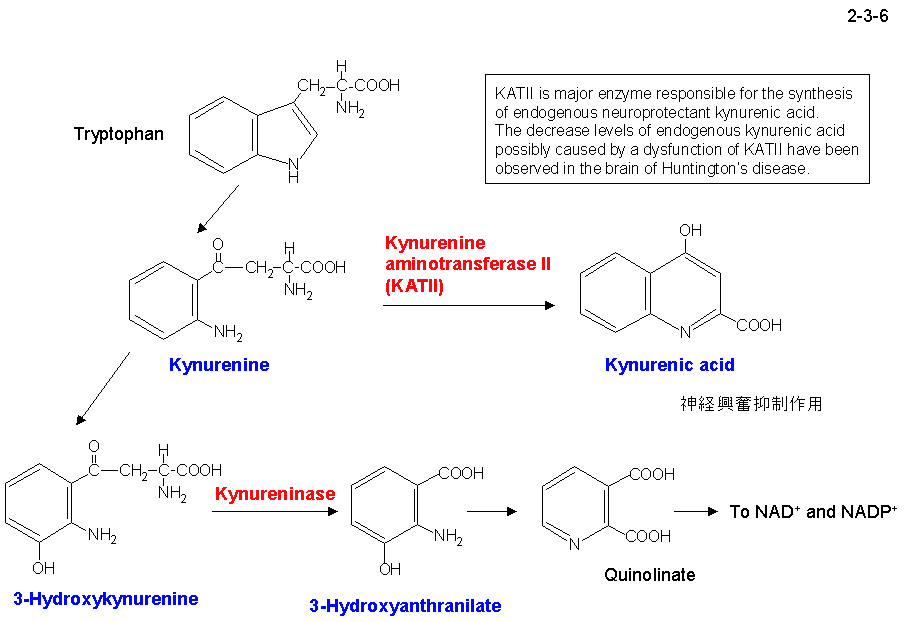
OT3 株由来キヌレニンアミノトランスフェラーゼ II(Ph-KAT II)は 428 アミノ酸残基から成り2量体として存在する。Ph-KAT II はヒト KAT II と 30% のアミノ酸配列の同一性を示す(2-3-5)。
KAT II はキヌレニンのアミノ基転移反応を触媒することによりキヌレニン酸を生成する(2-3-6)。KAT は一次構造の違いにより KAT I と KAT II の二つのファミリーに分類されるが、
KAT I と比べて KAT II の構造や機能の研究は遅れている。そこで、ヒト KAT II の基質認識機構や触媒機構を明らかにする目的で、そのホモログである Ph-KAT II の結晶化と構造解析をすすめている。
T. malitima由来 RNase HII(Tm-RNase HII)およびA. aeolicus由来RNases HII、HIII(Aa-RNases HII、HIII)はいずれも Tk-RNase HII 同様 type 2 RNase H である。しかし、
これらの酵素は超好熱性細菌由来という点で、超好熱始原菌(古細菌)由来の Tk-RNase HII とは異なる。前述のように、Tk-RNase HII は高度好熱菌由来 RNase HI(Tt-RNase HI)や大腸菌由来 RNase HI(Ec-RNase HI)より平衡論的にも速度論的にも安定化していることを明らかにしている。
しかし、RNase HI は type 1 RNase H であり、一次構造的にはRNase HII と大きく異なる。Tm-RNase HII や Aa-RNases HII と Tk-RNase HII の安定性を比較することにより、
同じファミリーの蛋白質同士でも超好熱細菌由来蛋白質と超好熱始原菌(古細菌)由来蛋白質では安定化機構が異なるかどうかを明らかにできる。また、T. malitima由来 RNase HI(Tm-RNase HI)は type 1 RNase H であるが、
N 末端に機能未知の付加配列をもつ。現在、これらの超好熱性細菌由来各種 RNase H の耐熱化機構を明らかにする目的で、これらの蛋白質の大量生産、精製、諸特性の解析、結晶化などを行っている。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}