(6)ファミリー I.3 リパーゼの分子機能に関する研究
ファミリー I.3 リパーゼは C 末端領域に分泌シグナルを持ち1ステップで菌体外に分泌されるという点で他のリパーゼと異なる(1-6-1)[Angkawidjaja,
C. & Kanaya, S. (2006) Cell. Mol.
Life Sci. 63, 2804-2817]。
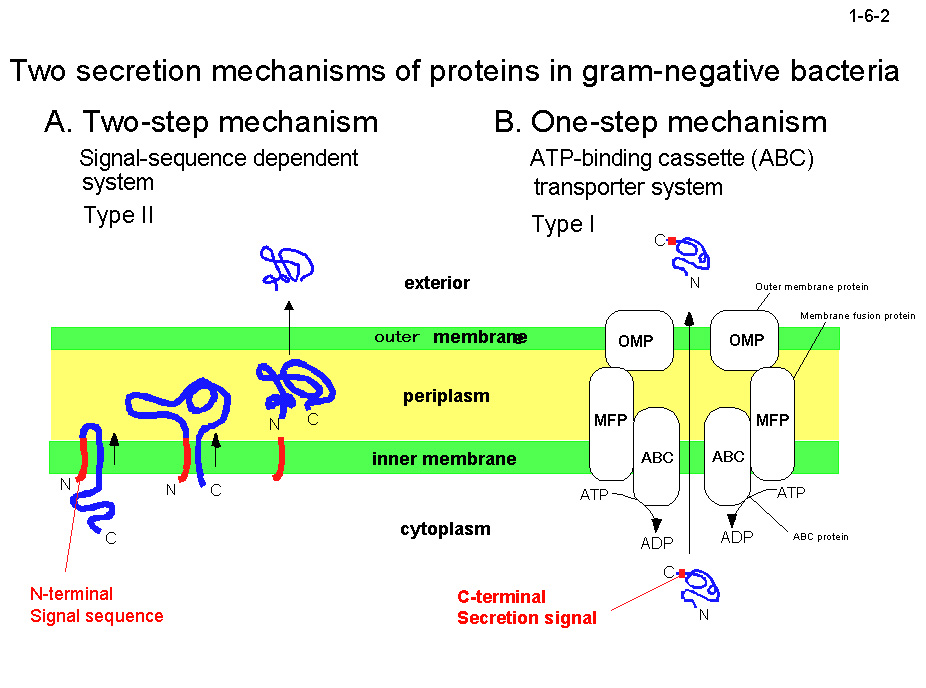
ファミリー I.3 リパーゼの分泌装置(Lipシステム)は ABC トランスポータあるいは I 型分泌システム(type1 secretion system, T1SS)とよばれ、3種類の蛋白質から構成される(1-6-2)。 このうち2つは ATP-binding cassette (ABC) protein と membrane fusion protein (MFP)とよばれる内膜蛋白質で、もう1つは outer membrane protein (OMP)とよばれる外膜蛋白質である。 ファミリー I.3 リパーゼ以外にも、ヘモリシン、リューコトキシンなどの毒素、メタロプロテアーゼ、ヘム結合蛋白質などがこのシステムにより分泌される。これらの蛋白質は分泌シグナルをN末端ではなく C 末端にもつ。 また、これらの蛋白質のC末端付近には通常GGXGXDXUX(X,
不特定残基; U, かさ高い疎水性残基)の9残基配列モチーフが繰返し存在している(1-6-3)。 この繰返し配列は Ca2+が結合するとβロール構造を形成すると考えられている(1-6-4)。
私達は、ファミリー I.3 リパーゼの構造機能相関や ABC トランスポータによる蛋白質の分泌機構を明らかにする目的で、Pseudomonas sp. MIS38 株由来リパーゼ(PML)の構造や機能、 Ca2+の役割、ドメイン構造の役割、分泌シグナルの役割などについて解析している。PML は 617 アミノ酸から成りモノマーとして存在する(1-6-5)[Amada,
K. et al. (2000) Biochim. Biophys. Acta
1478, 201-210]。 そのアミノ酸配列はS. marcescens
SM6 株由来リパーゼ (SML)と60.8%、P. fluorescens
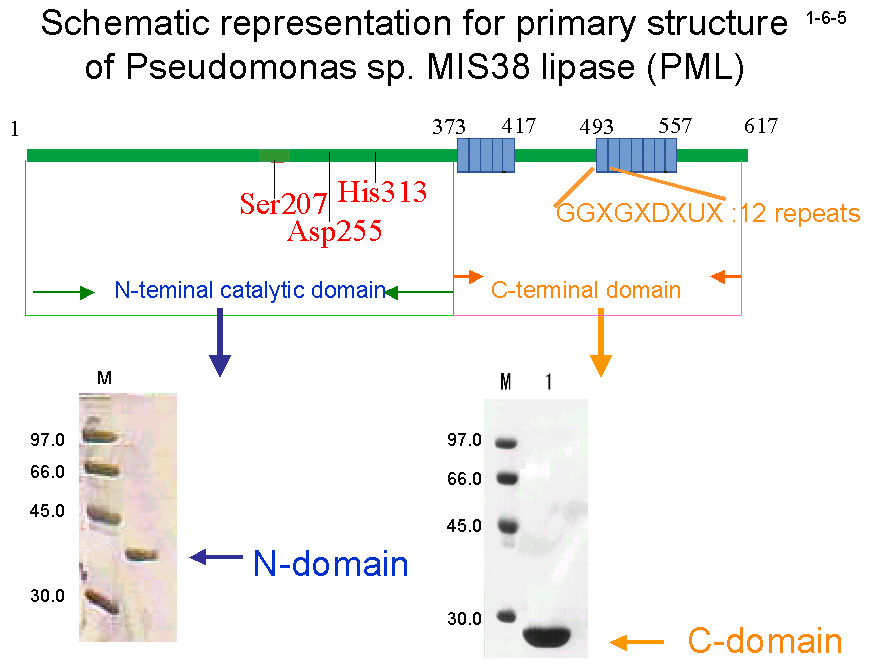
strain SIK W1 株由来リパーゼ(PFL)と 57.4 %の同一性を示す。PMLの活性部位は Ser207、Asp255、His313 により構成される[Kwon, H-J. et al. (2000) FEBS Lett. 483, 139-142]。
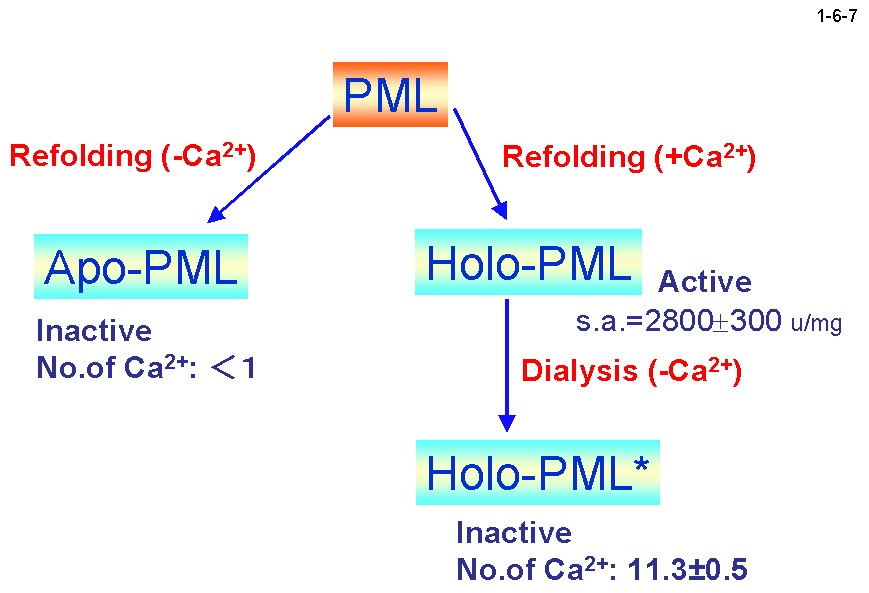
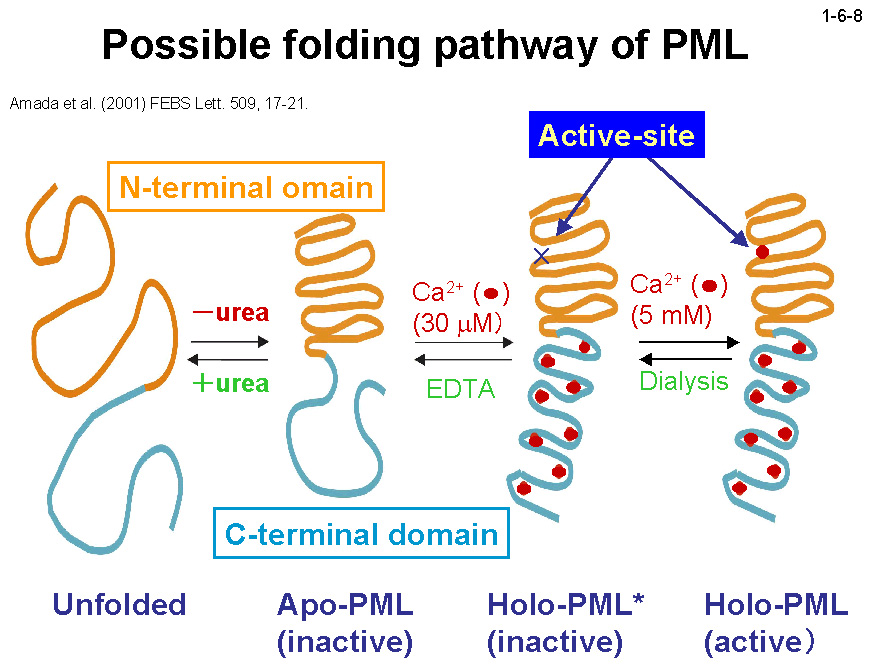
PML は触媒機構に関与する N ドメイン(1-372)と、分泌シグナル並びに 13 個の繰返し配列を含む C ドメイン(373-617)の2つのドメインに大きく分けられる。 PML は大腸菌で大量生産させるとインクルージョンボディとして不溶性画分に存在するため、精製する時は 8M 尿素存在下で行い、透析で尿素を除去することによりリフォールディングさせる(1-6-6)。 この時 Ca2+が存在すると Holo-PML が、存在しないと Apo-PML が生成する(1-6-7)。Apo-PML においては C ドメインはランダムコイル状態にあると考えられる(1-6-8)[Amada, K. et al. (2001) FEBS Lett. 509, 17-21]。
N ドメインはすべての活性残基を含むのにも関わらず単独では活性を示さないことから、活性部位が機能的なコンホメーションを形成するためには C ドメインが必要と考えられる[Kwon, H-J. et al. (2002) J. Biosci. Bioeng.
93, 157-164]。また、繰返し配列中の Asp を Ala に置換することによりβロール構造を形成しなくなった変異体 A387/A396-PML5 はほとんど培地中に分泌されず活性も示さないので、C ドメインのβロール構造は活性にも分泌にも大変重要と考えられる[Angkawidjaja,
C. et al. (2005) FEBS Lett. 579, 4707-4712]。
PML の分泌シグナルに関しては、C ドメインの繰返し配列を 13 回から 5 回まで減らしてもPMLの分泌量、酵素活性、安定性にはそれほど変化がないこと、 それ以上繰返し配列の数を減らすと蛋白質の分泌量や菌体内の蓄積量は著しく低下すること、などを明らかにしている(1-6-9)[Kwon,
H-J. et al. (2002) J. Biosci. Bioeng. 93,
157-164]。これらの結果は、繰返し配列が蛋白質の安定性に重要であることを示唆している。 また、C-末端から 19 残基を削ると PML は分泌されず活性も示さない。このことは、C-末端の 19 残基が分泌シグナルを含むこと、この領域は PML の分泌だけではなく PML が機能的なコンホメーションを形成するのに必要であることを示唆する。
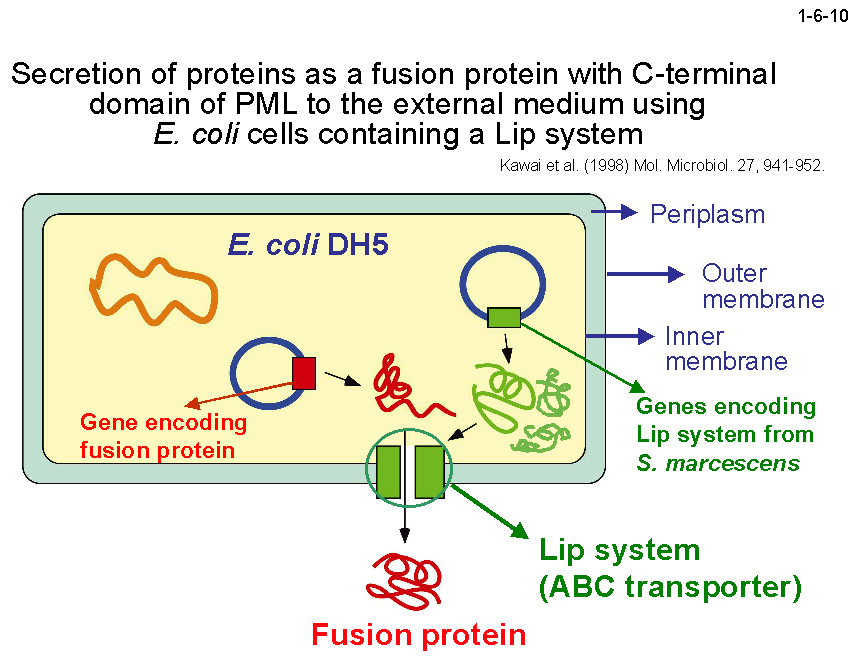
私達は、PML の C ドメインを利用して異種蛋白質を大腸菌の菌体外に分泌することも試みている(1-6-10)。蛋白質の構造解析はポストゲノム研究における重要な課題の一つであるが、
蛋白質の構造解析を行うためにはまず蛋白質の大量生産システムを構築する必要がある。大腸菌を用いて蛋白質を大量生産する場合、蛋白質を菌体内に蓄積させると高濃度になりすぎて凝集しやすくなり不溶化することがある。このような場合、
蛋白質を培地中に分泌させることができれば凝集による不溶化を防ぐことができるかもしれない。また、大腸菌のようなグラム陰性菌は蛋白質を培地中にはほとんど分泌しないので、大腸菌を用いて培地中に蛋白質を分泌させることができればその精製は極めて容易になる。
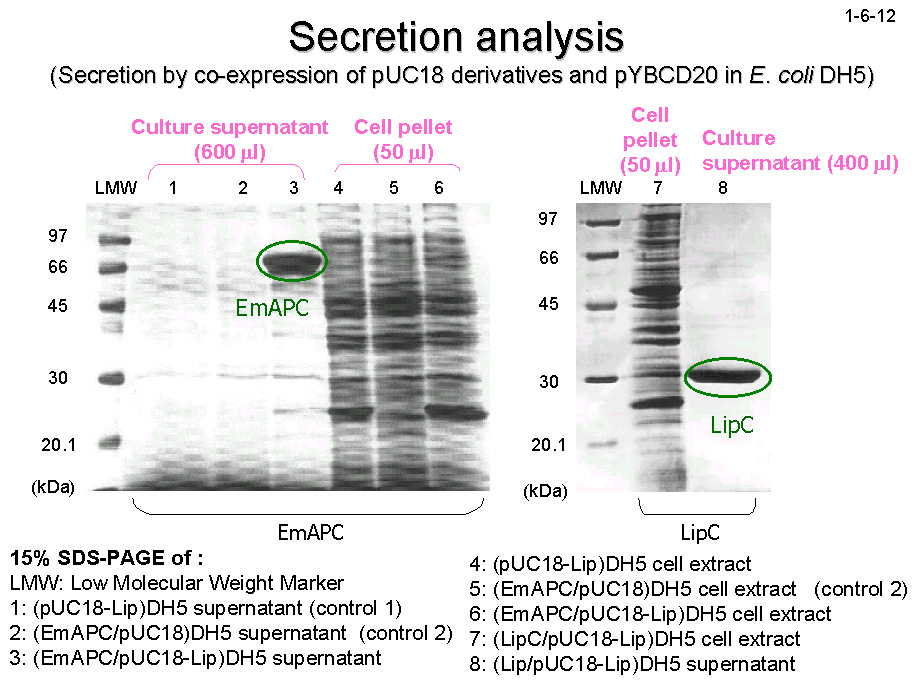
さらに大腸菌の場合は、培養が容易で生育速度も早く各種プラスミドも利用できるので蛋白質の大量生産に適している。私達はこれまでに、Lip システムを導入した大腸菌を宿主として PML(LipC)およびペリプラズム蛋白質である大腸菌アルカリホスファターゼの C 末端に LipC を連結した融合蛋白質を発現させると、
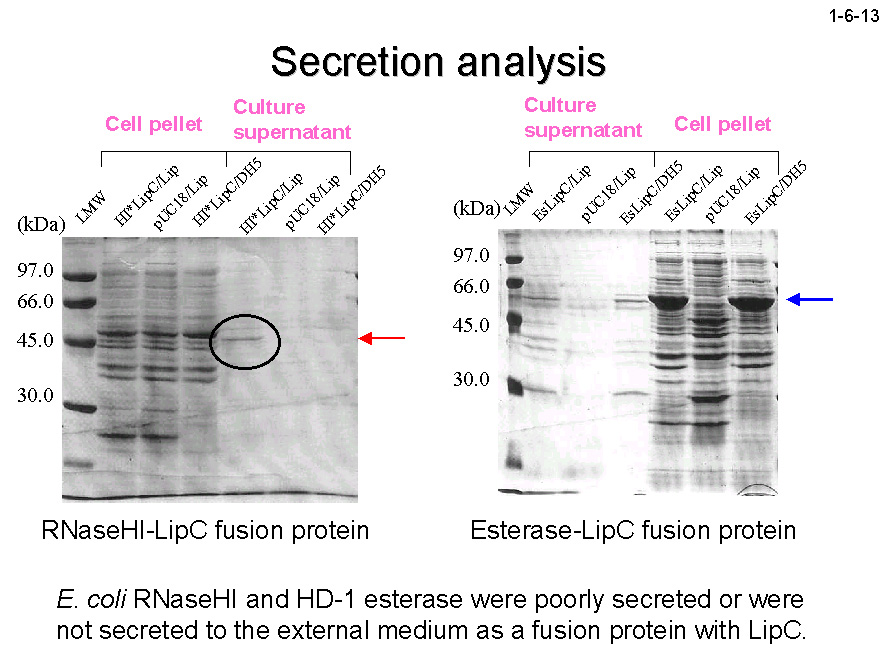
いずれも培地中に効率よく分泌されること(1-6-11、1-6-12)、一方、細胞質蛋白質である大腸菌 RNase HI やエステラーゼは PML は LipC を連結しても分泌されないこと(1-6-13)を明らかにしている[Angkawidjaja, C. et al.
(2006) Protein Eng. Des. Sel. 19, 337-343]。 細胞質蛋白質は合成されるとすみやかに折り畳まれるために分泌システムの狭いチャネルを通り抜けることができないのに対して、分泌蛋白質はゆっくり折り畳まれるために部分変性した状態で分泌システムを通り抜けることができるのかもしれない。
<2007年以後の研究>
1)PMLのlidが閉じた構造(closed構造)をX線結晶解析により決定した。同時期に決定されたSMLのlidが開いた構造(open構造)[Meier, R. et al. (2007) J. Biol. Chem. 282, 31477-31483]と比較することにより、ファミリー I.3 リパーゼは、他のリパーゼに保存されているlid1以外にlid2をもつこと、lid1が開いた構造はカルシウムイオンにより安定化されることを明らかにした[Angkawidjaja, C. et al. (2007)
FEBS Lett. 581,
5060-5064]。また、PMLのC末端ドメインはSML同様2つのβロール構造が並ぶβサンドイッチ構造を形成すること、1つ目のβロール構造は5個の繰返し配列から成りカルシウムイオンを3個含むこと、2つ目のβロール構造は8個の繰返し配列から成りカルシウムイオンを5個含むことを明らかにした。
2)PMLのNドメイン(触媒ドメイン)に結合する3個のカルシウムイオンの役割を解析する目的で、各結合部位(Ca1-Ca3サイト)に変異を導入し、活性や安定性に及ぼす影響を解析した。その結果、Ca1は安定性には関与しないが活性に必須であること、Ca2は活性に必須ではないが活性の向上と安定化に寄与すること、Ca3は活性に必須ではないが安定化に寄与することを明らかにした[Kuwahara, K. et al. (2008) Protein Eng. Des. Sel. 21, 737-744]。以前、PMLは1個のカルシウムイオンを活性に必要とし、その解離定数は0.13 mMであることを報告したが[Amada, K. et al. (2001) FEBS Lett. 509, 17-21]、本研究により、これはCa1であり、lid1を固定して不活性型のclosed構造から活性型のopen構造に変化させるのに必要であることが明らかになった。
3)PMLのopen構造をX線結晶解析により決定した。また、PMLのopen構造とclosed構造の結晶構造に基づいてMDシミュレーションを行うことにより、lidの開閉機構を解析した。その結果、closed構造からopen構造に変化する時はまずlid1が開くこと、open構造からclosed構造に変化する時はまずlid2から閉じることを明らかにした[Angkawidjaja, C. et al. (2009) J. Mol. Biol. 400, 82-95]。Ca1はlid1の固定化フックとして働き、lid1のopen構造を安定化しlid2が開くのを助けると考えられる。
4)PMLのC末端から15-19番目に存在する5残基配列モチーフ(VTLVG)とC末端に存在するDGIVIAモチーフはT1SSにより分泌されるタンパク質の間で良く保存されている。これらのモチーフの役割を明らかにする目的で、各種変異体を構築し、安定性、活性、分泌能を解析した。その結果、5残基配列モチーフは分泌に必須で、分泌シグナルとして働くこと、C末端モチーフは活性や分泌には必要ないが、安定化に寄与することを明らかにした[Kuwahara, K. et al. (2011) Protein Eng. Des. Sel. 24, 411-418]。
5)PMLのlid2の役割を明らかにする目的でlid2を欠損させた変異体を構築し、活性や安定性を解析した。その結果、lid2はinterfacial activationに必要であることを明らかにした[Cheng, M. et al. (2012) FEBS J. 279, 3727-3737]。また、lid2の欠損によりPMLのリパーゼ活性は低下し、エステラーゼ活性は向上したので、lid2はミセルを形成する基質への結合に必要であることが示唆された。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}