乮俆乯僺僩儕儔僀僔儞乮IDE 儂儌儘僌乯偺婎幙擣幆婡峔偵娭偡傞尋媶乮俀侽侽俇擭廔椆乯
戝挵嬠僺僩儕儔僀僔儞乮僾儘僥傾乕僛 Pi丄僾儘僥傾乕僛 III乯偼儊僞儘僾儘僥傾乕僛偱Zn2+丄Ca2+丄Co2+丄Mn2+側偳傪妶惈偵昁梫偲偡傞丅
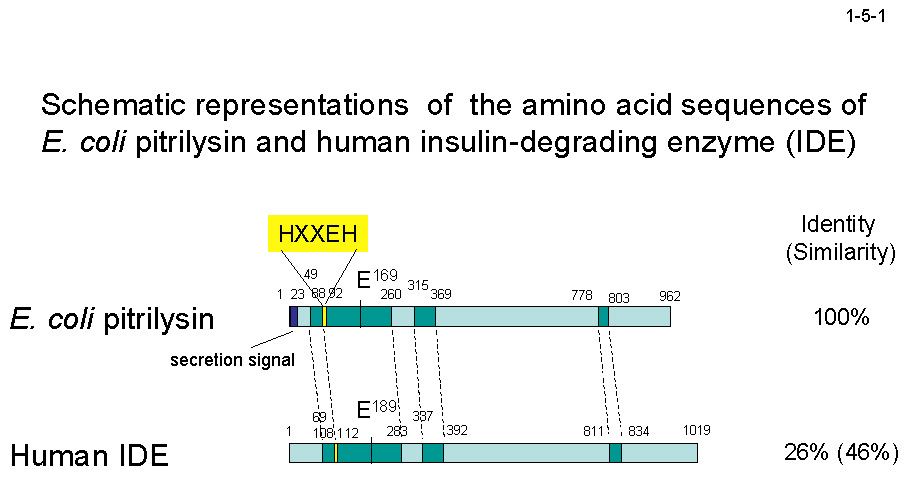
杮峺慺偼 962 傾儈僲巁巆婎偐傜惉傞慜嬱懱偲偟偰崌惉偝傟偨屻儁儕僾儔僘儉偵暘斿偝傟傞丅偦偺嵺丄N 枛抂偺 23 巆婎偐傜惉傞僔僌僫儖儁僾僠僪偼彍嫀偝傟傞偺偱丄杮峺慺偼 939 巆婎偐傜惉傞丅 杮峺慺偼儌僲儅乕偲偟偰摥偔丅杮峺慺偺傾儈僲巁攝楍偼丄歁擕椶桼棃僀儞僔儏儕儞暘夝峺慺乮Insulin-degrading enzyme, IDE乯偲 26 % 偺摨堦惈傪帵偡乮1-5-1乯丅傑偨丄N-arginine dibasic convertase丄
怉暔桼棃 Zn2+僾儘僥傾乕僛丄僸僩桼棃儊僞儘僾儘僥傾乕僛 I乮MPI乯丄僋儘儘僾儔僗僩僾儘僙僔儞僌峺慺乮CPE乯丄儅儔儕傾尨拵桼棃僼傽儖僔儕僔儞丄峺曣桼棃 AXL1 僾儘僥傾乕僛丄側偳偲傕 20-30 % 偺摨堦惈傪帵偡丅 偙傟傜偺峺慺偼僀儞僔儏儕僔儞僼傽儈儕乕傑偨偼儊僞儘儁僾僠僟乕僛 clan ME 僼傽儈儕乕 M16 偵暘椶偝傟傞丅
IDE偼僸僩偺懱撪偱偼擼傪偼偠傔偲偟偰條乆側慻怐偱敪尰偟偰偍傝丄僀儞僔儏儕儞丄beta僄儞僪儖僼傿儞丄惉挿場巕椶側偳條乆側儁僾僠僪儂儖儌儞椶傪攝楍摿堎揑偵暘夝偡傞丅
IDE 偼抈敀幙傗掅暘巕検偺崌惉婎幙傪暘夝偟側偄偺偱婎幙偺僒僀僘傪擣幆偡傞偲峫偊傜傟偰偄傞丅偟偐偟丄奺庬婎幙偺愗抐晹埵傪斾妑偟偰傕偦偺晅嬤偺傾儈僲巁攝楍偼曐懚偝傟偰偄側偄偨傔丄IDE 偑偳偺傛偆偵偟偰婎幙傪擣幆偡傞偺偐偼傢偐偭偰偄側偄丅 傑偨丄IDE 偺惗棟婡擻偵偮偄偰傕傛偔傢偐偭偰偄側偄丅偟偐偟嵟嬤丄IDE 偼傾儈儘僀僪儁僾僠僪傕暘夝偡傞偙偲偑尒偄偩偝傟偨乮1-5-2乯丅偙偺偙偲偼丄IDE 偑傾儈儘僀僪儁僾僠僪偺僗僇儀儞僕儍乕偲偟偰摥偔偙偲傪帵嵈偡傞丅
傾儈儘僀僪偺宍惉偼傾儖僣僴僀儅乕昦偺尨場偲峫偊傜傟偰偄傞偺偱丄IDE 偺婡擻傪憹嫮偡傞曽朄偼傾儖僣僴僀儅乕昦偺帯椕朄偲偟偰桳岠偐傕偟傟側偄丅
戝挵嬠僺僩儕儔僀僔儞傪戝検惗嶻偝偣傞偲杮峺慺偼儁儕僾儔僘儉偵拁愊偡傞丅1 儕僢僩儖攟梴塼偁偨傝偺惗嶻検偼 50-60 mg丄惛惢廂検偼 30-40 mg 偱偁傞丅杮峺慺偼抈敀幙傗掅暘巕検偺崌惉婎幙傪暘夝偟側偄偑丄 IDE 摨條僀儞僔儏儕儞丄beta 僄儞僪儖僼傿儞丄惉挿場巕椶側偳條乆側儁僾僠僪儂儖儌儞傪尷傜傟偨埵抲偱偺傒愗抐偡傞乮1-5-3乯丅傑偨丄beta-傾儈儘僀僪乮1-40乯傪 His14 偲 Gln15 偺娫偱摿堎揑偵愗抐偡傞乮1-5-4乯丅 偟偐偟丄奺儁僾僠僪婎幙偺愗抐揰晅嬤偺傾儈僲巁攝楍偵偼曐懚惈偼尒傜傟側偄丅偙傟傜偺寢壥偼丄僺僩儕儔僀僔儞偑儁僾僠僪婎幙偺崅師峔憿傪擣幆偡傞偙偲傪帵嵈偡傞[Cornista, J. et al. (2005) Biosci. Biotechnol. Biochem. 68, 2128-2137]丅儁僾僠僪偼悈梟塼拞偱偼儔儞僟儉僐僀儖峔憿傪傕偮偺偱丄
偙偺峔憿偼峺慺婎幙暋崌懱偑宍惉偝傟傞帪偵桿摫偝傟傞傕偺偲巚傢傟傞乮1-5-5乯丅僺僩儕儔僀僔儞偺妶惈拞怱偼 His 偲 Glu 偱峔惉偝傟偰偍傝丄
曐懚攝楍 HXXEH(X)76E乮88-169乯撪偺 His88 偲 His92 偲 Glu169 偼 Zn2+寢崌晹埵傪丄Glu91 偼怗攠晹埵傪宍惉偡傞乮1-5-1乯丅傑偨丄 兝傾儈儘僀僪儁僾僠僪傪戝挵嬠 RNase H1 偲偺梈崌抈敀幙偲偟偰戝検惗嶻偡傞僔僗僥儉傕峔抸偟偨[Cornista, J. et al. (2006) J. Biotechnol. 122,
186-197]丅偙偺僔僗僥儉傪梡偄傟偽儕僔儖僄儞僪儁僾僠僟乕僛偵傛傝梈崌抈敀幙偐傜beta傾儈儘僀僪儁僾僠僪傪愗傝弌偡偙偲偑偱偒傞偺偱丄
條乆側beta傾儈儘僀僪儁僾僠僪桿摫懱傪嶌惉偟僺僩儕儔僀僔儞偵傛傞暘夝條幃傪夝愅偡傞偙偲偑壜擻偱偁傞丅
丂側偍丄杮尋媶偼俀侽侽俇擭偵廔椆偟偨丅
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}