(3)大腸菌 RNase HI の構造安定性に関する研究
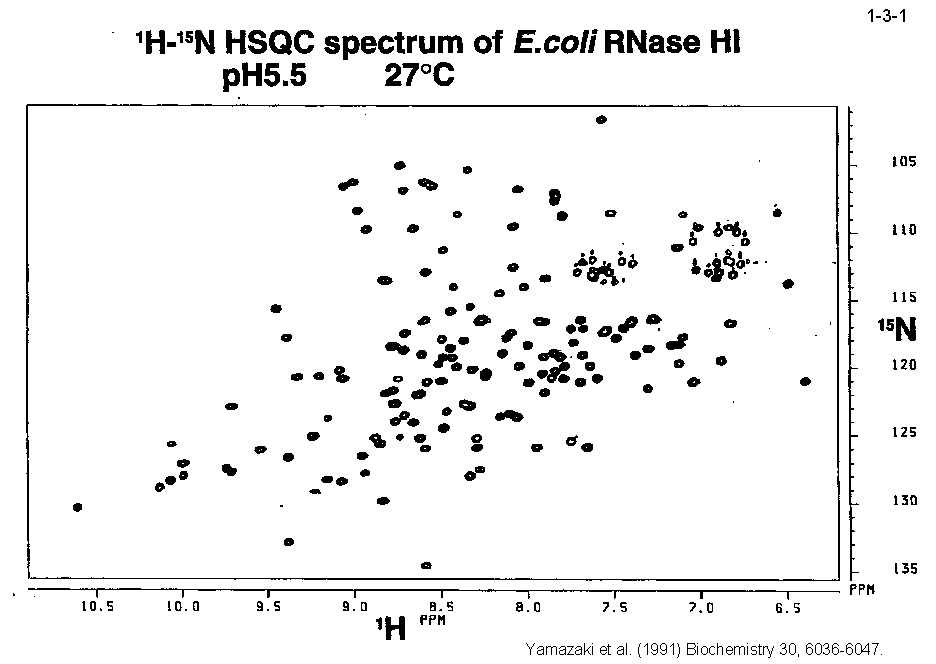
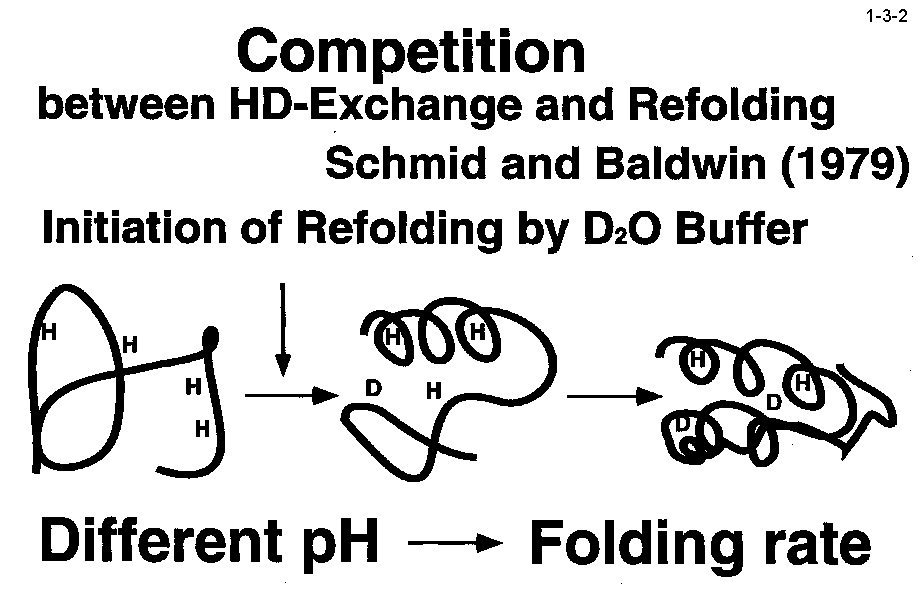
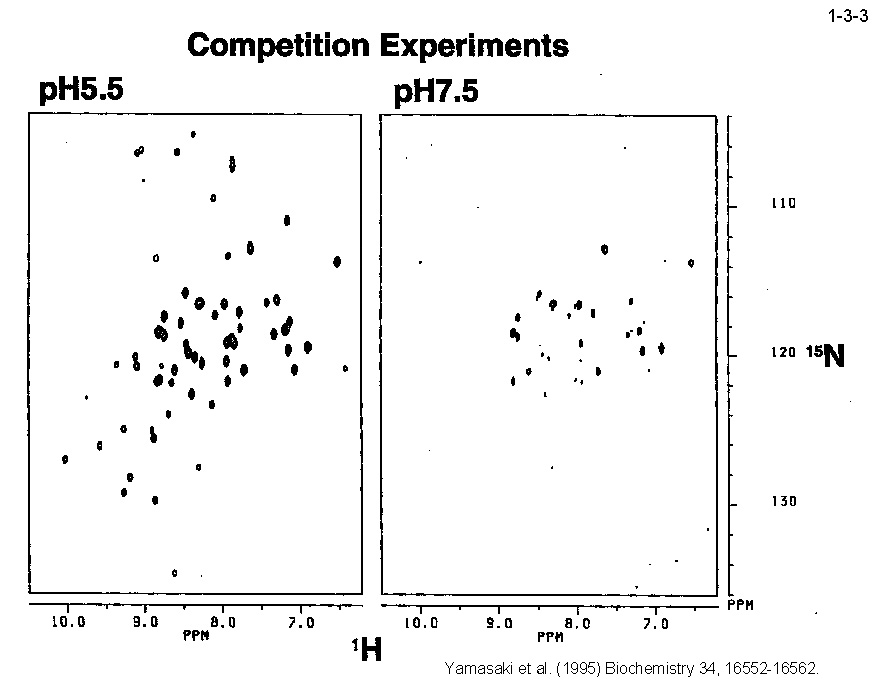
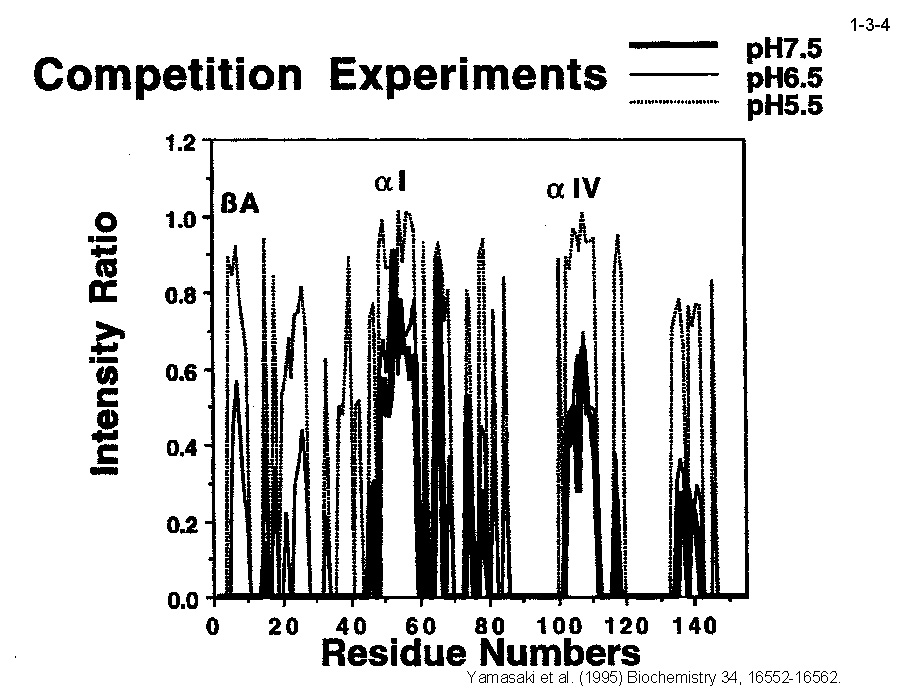
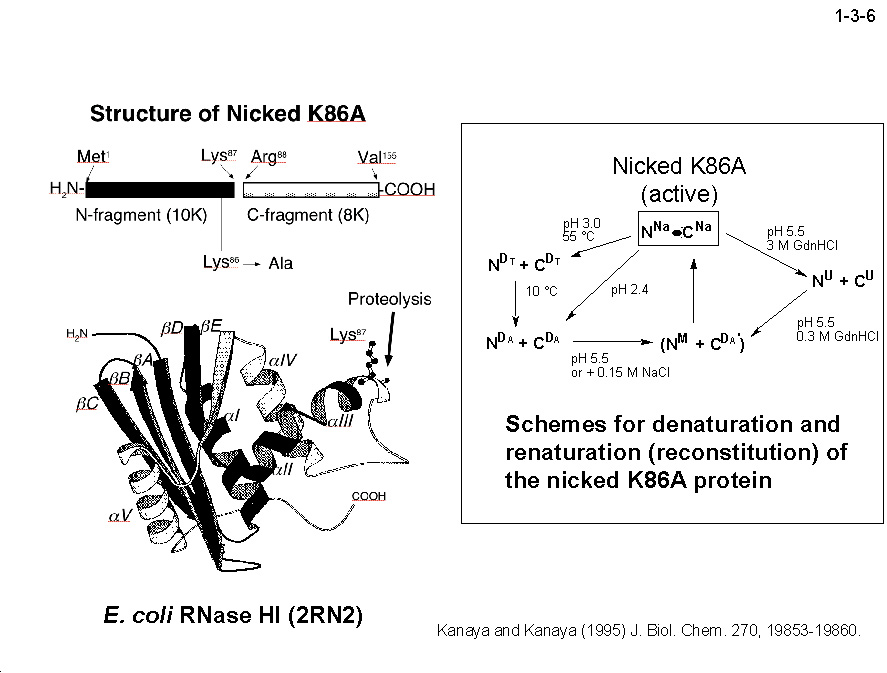
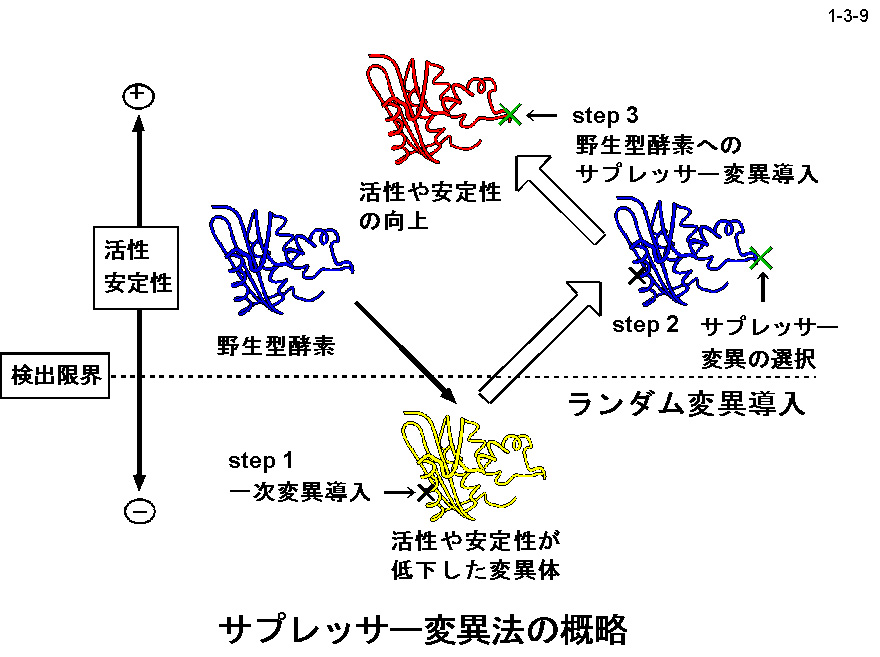
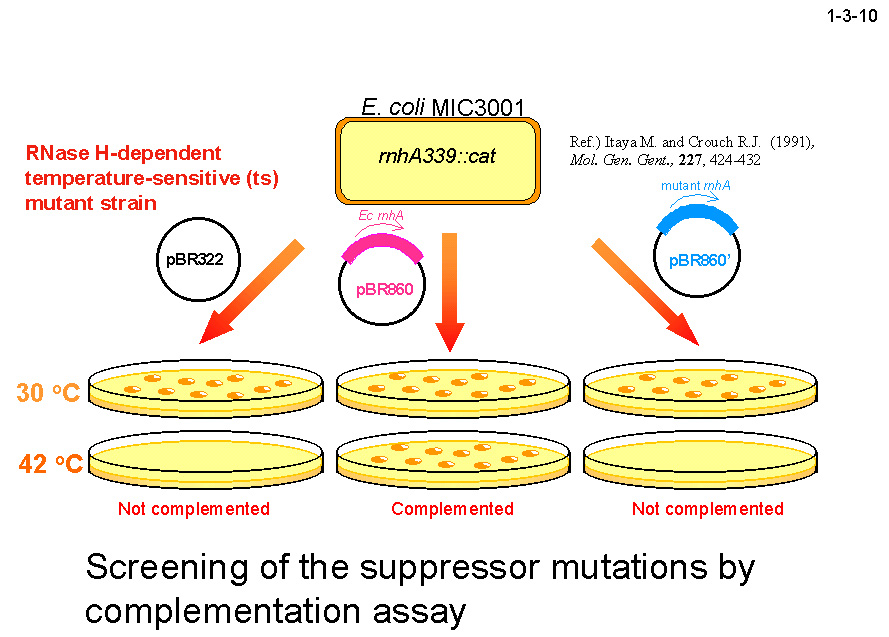
私達は、大腸菌 RNase HI をモデルとして蛋白質の安定化機構やフォールディング機構の解析も行っている。大腸菌 RNase HI は、 1)分子量が比較的小さい(19 kDa)、2)モノマーとして存在する、3)熱処理や変性剤処理に対して可逆的に変性する、4)変異体の構築および大量精製が容易である、 5)結晶化や構造解析が容易である、などの理由により蛋白質の構造ー安定性ーフォールディング相関の解析に適している。本酵素のフォールディングに関しては、 主鎖のアミドプロトンの交換反応とリフォールディング反応の競合を NMR で解析したり(1-3-1:大腸菌 RNase HI の2次元 NMR スペクトル; 1-3-2:実験方法;1-3-3:実験結果1;1-3-4:実験結果2;1-3-5:フォールディング反応の早い領域と遅い領域の分布)、 蛋白質を N 断片と C 断片に断片化して再構成したり(1-3-6)することにより、αヘリックス I がフォールディングの核になることを明らかにしている。 また本酵素の安定化に関しては、様々な変異体の安定性を野生型酵素と比較することにより(1-3-7)、ジスルフィド結合の導入、分子内疎水性コアの空孔充填、 ループ構造部分へのプロリン残基の導入、静電的反発の解消、左巻きヘリックス構造を形成するアミノ酸残基のグリシンへの置換、などにより本蛋白質の安定性が向上することを見いだしている(1-3-8)。 また、進化工学的な手法による本蛋白質の安定化も試みている(1-3-9)。RNase HI 遺伝子を破壊した大腸菌変異株 MIC3001 は温度感受性の性質を示し 42℃ では生育しないが、 RNase HI 遺伝子を導入すると42℃でも生育する(1-3-10)。しかし、C 末端の 13 残基を欠失させた変異体 142-RNas HI をコードする変異遺伝子を導入した MIC3001 株は 42℃ で生育しない。 142-RNas HI は全長蛋白質と比べると非常に不安定で、42℃ では変性してしまうためである。従って、この変異遺伝子にランダム変異を導入し、MIC3001 株を形質転換した後、 42℃ でも生育できる形質転換体をスクリーニングすれば、このような形質転換体は 142-RNas HI の安定化変異体をコードする遺伝子(サプレッサー変異遺伝子)をもつことが期待される。 ついでこの変異体の安定化に寄与するアミノ酸置換を同定し、全長蛋白質に導入すれば、このアミノ酸置換が全長蛋白質も同様に安定化するかどうかがわかる。このようにして、 これまでに 10 種類の安定化変異を同定することに成功している(1-3-11)。部位特異的変異法や進化工学的手法により得られた大腸菌 RNase HI 変異体の構造と安定性を系統的、 網羅的に解析することにより、蛋白質の安定化機構、さらには構造構築機構に関して有用な情報が得られると期待される。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}