(2)RNase H の分子多様性に関する研究
RNase H は、岡崎フラグメントの合成に必要なRNAプライマーの生成や分解除去、転写の過程で生成する不必要な R-loop の分解除去、
プラスミドや染色体の特異的な複製開始点以外からの複製開始の阻害、などに関与すると考えられている。また、DNAの複製中に間違って取り込まれた RNA の除去にも関与すると考えられている。
RNase H はその一次構造の違いにより Type 1 RNase H と Type 2 RNase H の2つのファミリーに大きく分類される(1-2-1)。
原核生物由来酵素は RNases HI、HII、HIII の3つのグループに分けられるが、RNase HI は Type 1 に、RNase HII と RNase HIII は Type 2 に属する。
また、真核生物由来酵素は RNase H1 と RNase H2 の2つのグループに分けられるが、 RNase H1 は Type 1 に、RNase H2 は Type 2 に属する。
逆転写酵素の RNase H ドメインは Type 1 に属する。Type 1 RNase H と Type 2 RNase H の一次構造を比べると相同性を示す領域はほとんどないので、
これらの酵素は進化的に異なる祖先をもつと考えられる(1-2-2)。従って、Type 1 RNase H と Type 2 RNase H が共通の三次構造モチーフ(RNase H-fold)をもつのは、
収束進化によると思われる。
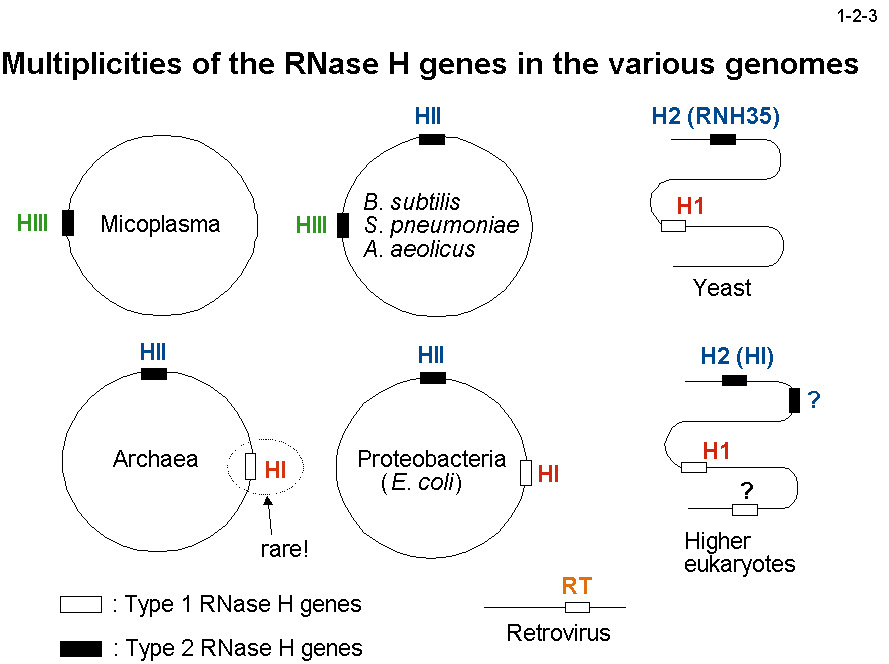
RNase H 遺伝子は一つのゲノム上に通常2つある(1-2-3)。しかし、その種類は生物によって異なる。例えば、大腸菌のゲノムは RNase HI と RNase HII の遺伝子をもち、
枯草菌のゲノムは RNase HII と RNase HIII の遺伝子をもつ。酵母やヒトのゲノムは RNase H1 と RNase H2 の遺伝子をもつ。また、主要な活性を担う RNase H も生物によって異なる。
例えば、低温菌Shewanella sp. SIB1 株の場合は RNase HII が、大腸菌の場合は RNase HI が、枯草菌の場合は RNase HIII が主要な活性を担う。大腸菌、枯草菌、酵母の RNase H 遺伝子破壊実験の結果、
1)2つのrnh 遺伝子は細胞内で両方とも機能している、2)一方の遺伝子を破壊しても他方の遺伝子がその機能を補う、3)RNase H 活性は細胞の正常な生育に必要である、ことなどが示唆されている。
しかし、自然界には Type 1 か Type 2 どちらかの RNase H しかもたない生物も存在するので、生物の進化と RNase H の多様性には何らかの相関があると思われ興味深い。
私達は、生物の進化に関して新しい知見を得ることをめざして RNase H の分子多様性について解析している。
RNase H の基質結合部位の構造には多様性がみられる。例えば、大腸菌 RNase HI においては分子中央部にある塩基性突出部位が、超好熱始原菌(古細菌)RNase HII においては C 末端ドメインが、
枯草菌 RNase HII においてはN末端付加配列が、基質結合に重要である(1-2-4)。このことは、RNase H 分子が進化の過程で触媒ドメインと基質結合ドメインのシャフリングにより多様化してきたことを示唆しており興味深い。
私達は、N 末端領域もC末端領域も基質結合に重要な枯草菌 RNase HIII がどのような構造をもつのか、そしてどのようにして基質と結合するのかを明らかにする目的で、枯草菌 RNase HIII より安定で大量に精製しやすい
Bacillus stearothermophilus RNase HIII の結晶構造を解析している。
{kind=link}
{kind=link}
{kind=link}
{kind=link}